
Abstract
SARS-CoV-2 transmission is uncontrolled in many parts of the world, compounded in some areas by higher transmission potential of the B1.1.7 variant now seen in 50 countries. In addition, many countries are extending the interval between doses of vaccine is being extended in order to expand vaccine coverage. It is unclear whether real world neutralising antibody titres will follow those in clinical trials, particularly in elderly persons, and how this will be impacted by mutations in strains such as B1.1.7. Here we tested immune responses in patients 3 weeks following the first dose of the Pfizer BioNTech vaccine BNT162b2. We also tested neutralising antibody responses against pseudoviruses expressing wild type Spike proteins or expressing 3 key mutations present in B.1.1.7 (deletion 69/70, N501Y, A570D). IgG Spike antibody titres correlated well with neutralisation. We observed a range of neutralisation titres against wild type, with geometric mean titre (GMT) of 24. The vaccine sera exhibited a range of inhibitory dilutions giving 50% neutralisation (ID50) from <1:4 to 3449. However a lower proportion of participants over 80 years old achieved threshold neutralisation titre of >1:4 for 50% neutralisation as compared to those under 80 years old (8/15 versus 8/8 P=0.052) after the first dose. Neutralisation titres were not significantly impacted by the combination of three Spike mutations tested, but were reduced against the full set of Spike mutations present in the B.1.1.7 variant. The highest fold change was approxiamately 6 and the median fold change for the B.1.1.7 variant versus wild type was 3.85 (IQR 2.68-5.28). Further work is needed to establish the impact of these observations on real life vaccine efficacy.
Introduction
The outbreak of a pneumonia of unknown cause in Wuhan, China in December 2019, culminated in a global pandemic due to a novel viral pathogen, now known to be SARS-COV-21. The unprecedented scientific response to this global challenge has led to the rapid development of vaccines aimed at preventing SARS-COV-2 infection and transmission. Alongside these developments, the virus has continued to evolve to optimise its infectivity/transmissibility2,3,4,5 and to evade drug6 and immune control7.
SARS-CoV-2 vaccines have recently been licensed that target the Spike protein, either using mRNA or adenovirus vector technology with protection rates over a few months ranging from 62-95%8-10. The Pfizer BioNTech BNT162b2 vaccine encodes the full-length trimerised spike protein of SARS COV-2 and is formulated in lipid nanoparticles to optimise delivery to cells11. Other vaccines include the Moderna mRNA-1273 vaccine, also lipid nanoparticle formulated spike glycoprotein12. The Oxford-AstraZeneca ChAdOx1 nCoV-19 vaccine (AZD1222) is a replication-deficient chimpanzee adenoviral vector ChAdOx1, containing the spike glycoprotein13. The duration of immunity conferred by these vaccines is as yet unknown. These vaccines were designed against the Wuhan strain isolated in 2019. Concerns have been raised as to whether these vaccines will be effective against the new variants of SARS-COV-2 recently emergent in the UK, South Africa and Brazil 14-16.
The phase I/II studies of the Pfizer BioNTech BNT162b2 vaccine determined the immunogenicity of different dosing regimens. The geometric mean concentration (GMC) of RBD-binding IgG 21 days after the first dose of 30ug of the BNT162b2 vaccine which is the dose approved in the UK, was higher than the GMC of a panel of convalescent plasma (1,536 vs 602 U/ml). However corresponding neutralisation geometric mean titre (GMT) was 3-fold lower than the panel of convalescent plasma (29 vs 94)11. However, after the second vaccine dose was given these neutralisation titres substantially increased. In older adults mean GMT was only 12 in a small analysis of 12 participants17 and increased to 109 after the second dose.
There are no real world data on vaccine immune responses following the BNT162b2 vaccine and no data on neutralisation of the UK variant B.1.1.7, now set to expand in the US. In this study we assess the immunity induced three weeks after one dose of the BNT162b2 vaccine in individuals who have received the first dose following the rollout in the UK. Furthermore, we investigate if three mutations present in the UK variant B.1.1.7 confer altered neutralisation sensitivity to sera from individuals vaccinated with the BNT162b2 vaccine.
Results
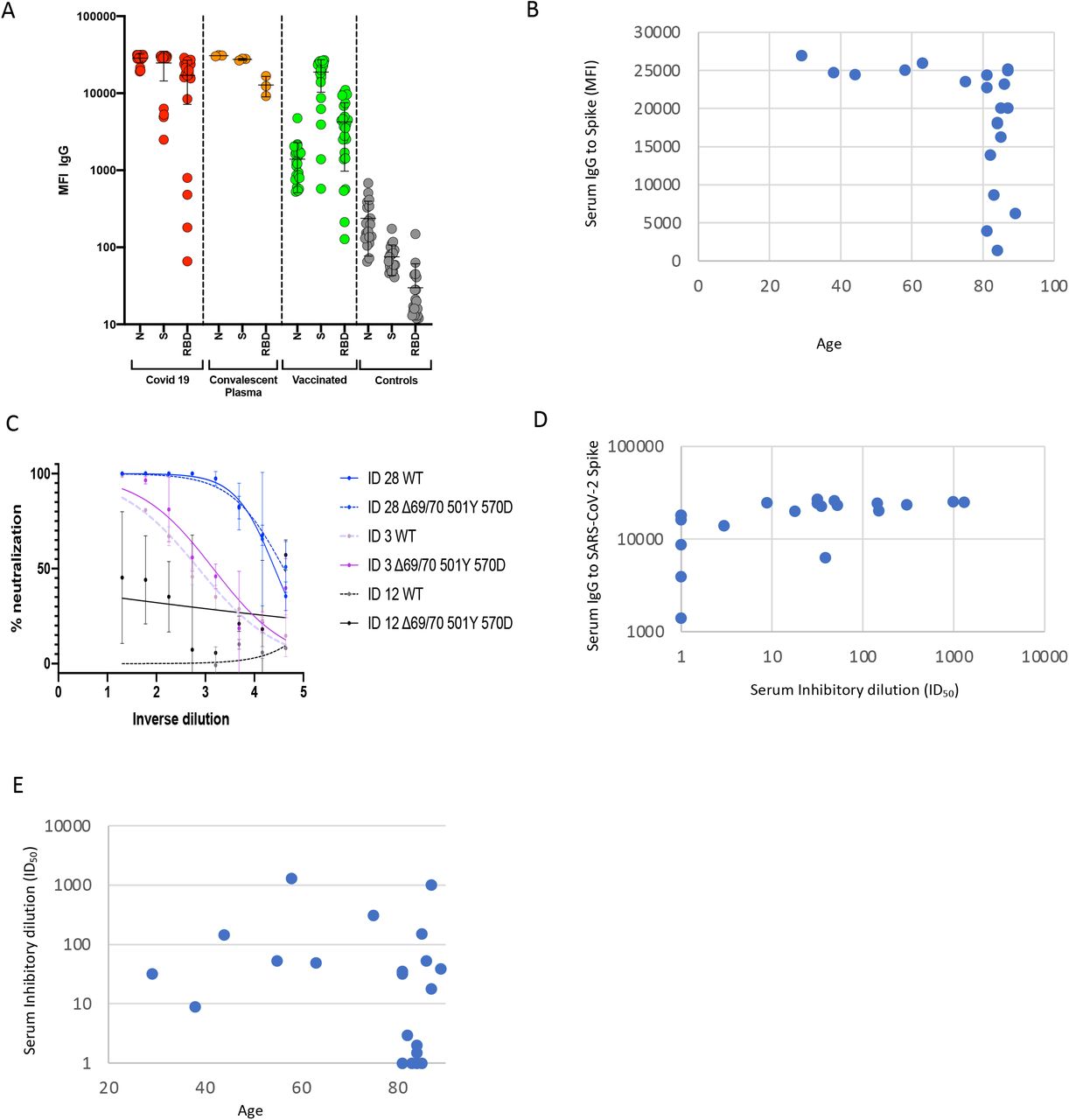
23 participants had received the vaccine three weeks prior to blood draw for serum and PBMC. Median age was 82 years (IQR 64-85) and 30% were female. Serum IgG titres to N protein, Spike and the Spike RBD were assayed by a Luminex bead system using flow cytometry (Figure 1A). These data showed titres much higher than in healthy controls, and similar to convalescent plasma and recovered individuals. However there was almost 100-fold variation in IgG responses to Spike and Spike RBD across the vaccinated participants. There appeared to be reduced IgG responses to Spike above the age of 80 (Figure 1B).

A. Serum IgG responses against N protein, Spike and the Spike Receptor Binding Domain (RBD) from 23 vaccinated participants (green), 20 recovered COVID-19 cases (red), 3 convalescent plasma units and 20 healthy controls (grey) as measured by a flow cytometry based Luminex assay. MFI – mean fluorescence intensity. B. Scatter plot of Serum IgG responses versus age of participants. C. Examples of three vaccinee participants’ data from serum pseudotyped virus Neutralisation assay. Indicated is serum dilution on the x-axis and % neutralisation on y-axis. These data are used to calculate the ID50 – the serum dilution required to inhibit 50% of virus infection (ID50). Data points represent means of technical replicates. D. Relationship between Serum IgG responses as measured by flow cytometry and serum neutralisation ID50. E. Relationship between ID50 and age.
Using lentiviral pseudotyping we generated wild type Spike proteins on the surface of enveloped virions in order to measure neutralisation activity of vaccine sera against these viruses. This system has been shown to give generally similar results to replication competent virus18,19. The vaccine sera exhibited a range of inhibitory dilutions giving 50% neutralisation (ID50) – <4 to 3449 (Figure 1C, D). The GMT against wild type (WT) pseudovirus was 24. Eight of 23 participants exhibited no appreciable neutralisation against the WT virus. There was reasonable correlation between full length Spike IgG titres and serum neutralisation (Figure 1, coefficient 0.37). Age was negatively correlated with serum neutralisation (Figure 1D; correlation coefficient -0.45). We observed poor neutralisation activity (<50% neutralisation at serum dilution of 1:4) exclusively in participants over the age of 80, Figure 1E, (7/15) as compared to those under 80 (0/8, p=0.05 by Fisher’s exact).
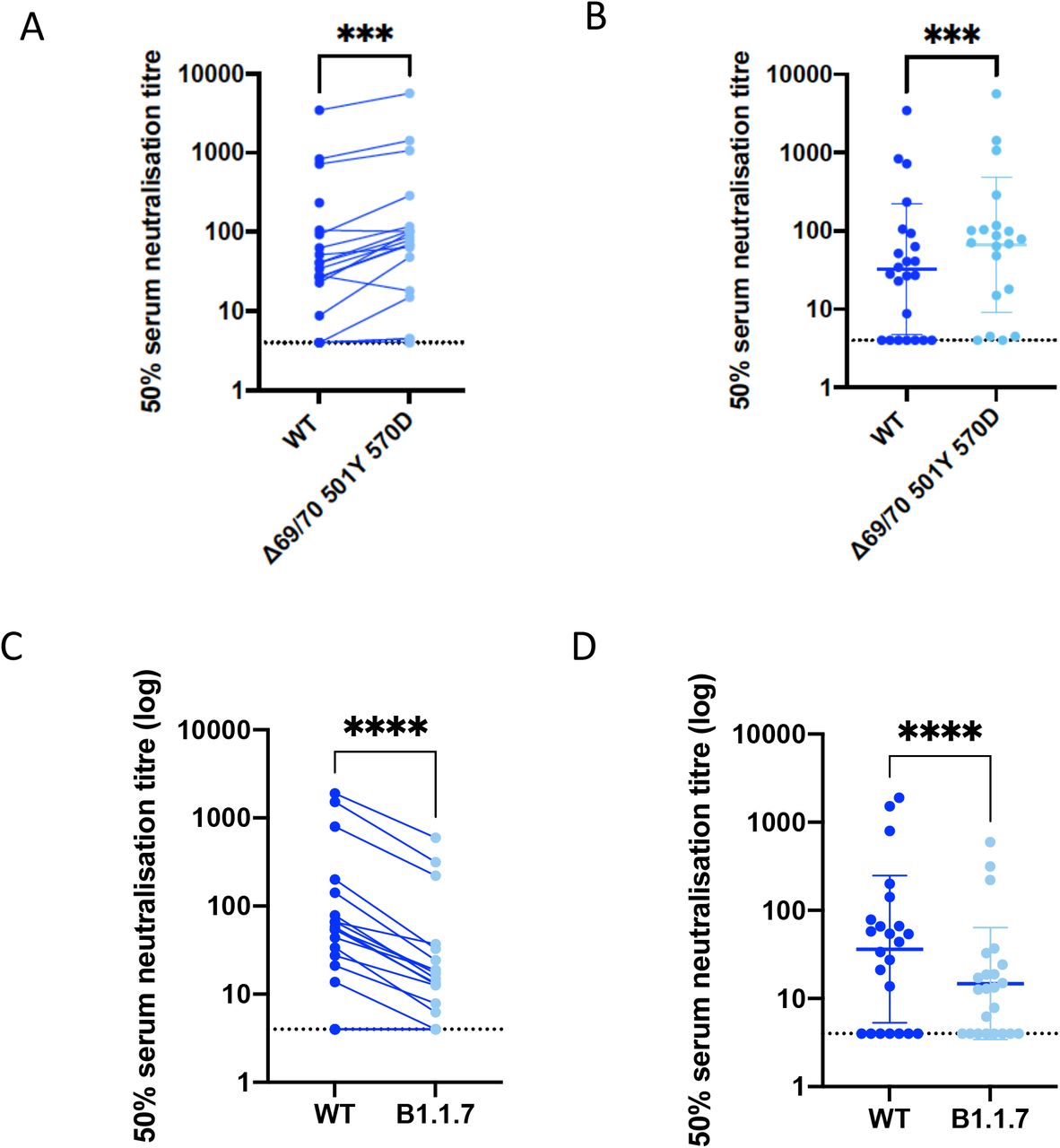
We next generated mutated Spike protein with mutations N501Y, A570D and the 69/70 deletion by site directed mutagenesis. We tested a panel of sera from eleven recovered individuals and found that these sera also neutralised both wild type and the mutated viruses similarly (Figure 2). We next tested this mutant Spike against wild type in a serum neutralisation assay using vaccinee sera. We observed no reduction in the ability of sera from vaccinees to inhibit either WT or mutant virus (Table 1, Figure 2).
Participants and serum neutralisation data; ID50 (inhibitory dilution) is the dilution required to achieve neutralisation of 50% infection by the virus. fold change (FC). NA: not available.

Indicated is serum log10 inverse dilution against % neutralisation. Where a curve is shifted to the right this indicates the virus is less sensitive to the neutralising antibodies in the serum. Data points represent means of technical replicates and error bars represent standard deviation.
We next generated the full set of eight mutations in the Spike protein (Figure 3), comprising RBD (N501Y and A570D), S1 (ΔH69/V70 and Δ144/145) and S2 (P681H, T716I, S982A and D1118H). We found that among 15 individuals with neutralisation activity three weeks after the Pfizer mRNA vaccine, 10 showed evidence of reduction in efficacy of antibodies against the B.1.1.7 mutant (Fold change >3, Figure 3, Table 1). The highest fold change was approxiamately 6 and the median fold change was 3.85 (IQR 2.68-5.28).
{kind=link}
{kind=link}
{kind=link}
N501Y, A570D and ΔH69/V70. C,D. B.1.1.7 contains N501Y, A570D, ΔH69/V70, Δ144/145, P681H, T716I, S982A and D1118H. Data are representative of 2 independent experiments in A, B.
Discussion
Amidst high transmission in many parts of the world, vaccines are a key part of a long term strategy to bring SARS-CoV-2 under control. Vaccine supply is limited, forcing some governments to consider extending the time between first and second doses. Information on vaccine elicited responses in real world settings outside of highly selective clinical trials are important, including studies in older people.
Serum neutralizing activity is a correlate of protection for other respiratory viruses, including influenza20 and respiratory syncytial virus where prohylaxis with monoclonal antibodies has been used in at risk groups21,22. Neutralising antibody titres appered to be highly correlated with vaccine protection against rechallenge in non human primates, and importantly, there was no correlation between T cell responses (as measured by ELISPOT) and protection23. Coupled with multiple reports of re-infection, there has been therefore significant attention placed on virus neutralisation.
The participants in this study had a median age of over 80 in line with the targeting of this age group in the initial rollout of the vaccines in the UK. Participants showed similar neutralising activity against wild type pseudovirus as in the phase I/II study, with geometric mean titres of 24 and 29 respectively11. In a study specifically looking at older adults the GMT after first dose was 12 in a set of 12 subjects between ages of 65 and 8517, rising to 109 seven days after the second dose. We also oberved lower neutralisation potency in participants over the age of 80 as compared to those under 80. This may or may not be compensated for by the second dose and it will be important to follow all participants over the following months for measurement of neutralisation activity as well as data on re-infection.
The three mutations in Spike (N501Y, A570D, del 69/70) did not appear to impact neutralisation in a pseudovirus assay. However we demonstrated that a pseudo virus bearing Spike protein with all Spike mutations in the B.1.1.7 variant (del69/70, del 144/145, N501Y, A570D, P681H, T716I, S982A, D1118H) did result in reduced sensitivity to sera from vaccinees. These modest changes are what we expected to see given the mutational profiles. Vaccines should still retain good activity and vaccine coverage is a priority.
Other variants also need to be tested against vaccine sera, some with more concerning mutations such as E484K and K417N that have been shown to impact neutralisation by monoclonal antibodies or convalescent sera15,24-26.
Limitations
Our study was limited by relatively small sample size. Although the Spike pseudotyping system has been shown to faithfully represent full length infectious virus, there may be determinants outside the Spike that influence antibody susceptibility either directly or indirectly in a live replication competent system. On the other hand live virus systems allow replication and therefore mutation to occur, and rigorous sequencing at multiple steps is needed. In addition, the cells used in such live virus systems are monkey cells engineered to overexpress receptors as in the case of our work. Finally, T cell responses are not accounted for in the present study.
Data Availability
Data are available from the corresponding author on request
Methods
Participant recruitment and ethics
Participants who had received the first dose of vaccine and individuals with COVID-19 were recruited into the NIHR Bioresource. The study was approved by the East of England – Cambridge Central Research Ethics Committee (17/EE/0025).
SARS-CoV-2 serology by multiplex particle-based flow cytometry (Luminex)
Recombinant SARS-CoV-2 N, S and RBD were covalently coupled to distinct carboxylated bead sets (Luminex; Netherlands) to form a 3-plex and analyzed as previously described (Xiong et al. 2020). Specific binding was reported as mean fluorescence intensities (MFI).
Generation of Spike mutants
Amino acid substitutions were introduced into the D614G pCDNA_SARS-CoV-2_Spike plasmid as previously described27 using the QuikChange Lightening Site-Directed Mutagenesis kit, following the manufacturer’s instructions (Agilent Technologies, Inc., Santa Clara, CA). Sequences were checked by Sanger sequencing.
Pseudotype virus preparation
Viral vectors were prepared by transfection of 293T cells by using Fugene HD transfection reagent (Promega). 293T cells were transfected with a mixture of 11ul of Fugene HD, 1µg of pCDNAΔ19Spike-HA, 1ug of p8.91 HIV-1 gag-pol expression vector28,29, and 1.5µg of pCSFLW (expressing the firefly luciferase reporter gene with the HIV-1 packaging signal). Viral supernatant was collected at 48 and 72h after transfection, filtered through 0.45um filter and stored at -80°C. The 50% tissue culture infectious dose (TCID50) of SARS-CoV-2 pseudovirus was determined using Steady-Glo Luciferase assay system (Promega).
Serum/plasma pseudotype neutralization assay
Spike pseudotype assays have been shown to have similar characteristics as neutralisation testing using fully infectious wild type SARS-CoV-218. Virus neutralisation assays were performed on 293T cell transiently transfected with ACE2 and TMPRSS2 using SARS-CoV-2 Spike pseudotyped virus expressing luciferase30. Pseudotyped virus was incubated with serial dilution of heat inactivated human serum samples or sera from vaccinees in duplicate for 1h at 37°C. Virus and cell only controls were also included. Then, freshly trypsinized 293T ACE2/TMPRSS2 expressing cells were added to each well. Following 48h incubation in a 5% CO2 environment at 37°C, luminescence was measured using the Steady-Glo Luciferase assay system (Promega).
Author contributions
Conceived study: RKG, DAC.
Designed study and experiments: RKG, DAC, LEM, JB, MW, JT, LCG, GBM, RD, BG, NK, AE
Performed experiments: BM, DAC, RD, IATMF, LCG, GBM,
Interpreted data: RKG, DAC, BM, RD, IATMF, LEM, JB, KGCS,
Acknowledgements
We would like to thank Cambridge University Hospitals NHS Trust Occupational Health Department. We would also like to thank the NIHR Cambridge Clinical Research Facility and staff at CUH and. We thank Dr James Voss for the kind gift of HeLa cells stably expressing ACE2. RKG is supported by a Wellcome Trust Senior Fellowship in Clinical Science (WT108082AIA). LEM is supported by a Medical Research Council Career Development Award (MR/R008698/1). SAK is supported by the Bill and Melinda Gates Foundation via PANGEA grant: OPP1175094. DAC is supported by a Wellcome Trust Clinical PhD Research Fellowship. This research was supported by the National Institute for Health Research (NIHR) Cambridge Biomedical Research Centre, the Cambridge Clinical Trials Unit (CCTU). JAGB is supported by the Medical Research Council (MC_UP_1201/16).
References




