
Abstract
In the last decade, despite the proven efficacy of vaccines, the developed world has seen a resurgence of vaccine-preventable diseases (VPDs) such as measles, pertussis, and polio. Vaccine hesitancy, an individual behavior influenced by historical, political, and socio-cultural factors, is believed to be a primary factor responsible for decreasing vaccine coverage, thereby increasing the risk and occurrence of VPD outbreaks. Society, culture, and individual motivations affect human decisions regarding health behavior and preventative care, and health perceptions and health-related behaviors can change at the population level as cultures evolve. In recent years, mathematical models of disease dynamics have begun to incorporate aspects of human behavior, however they do not address how evolving cultures influence these health behaviors. Here, using a mathematical modeling framework, we explore the effects of cultural evolution on vaccine hesitancy and vaccination behavior. With this model, we shed light on the facets of cultural evolution (vertical and oblique transmission, homophily, etc.) that promote the spread of vaccine hesitancy, ultimately affecting levels of vaccination coverage and VPD outbreak risk in a population. In addition, we present our model as a generalizable framework for exploring cultural evolution when beliefs influence, but do not strictly dictate, human behaviors. We show vaccine confidence and vaccine-conferred benefits can be driving forces of vaccine coverage, and we demonstrate that an assortative preference among vaccine-hesitant individuals can lead to increased vaccine hesitancy and lower vaccine coverage. Further, we show that vaccine mandates can lead to a phenomenon in which high vaccine hesitancy co-occurs with high vaccination coverage, and that high vaccine confidence can be maintained even in areas where access to vaccines is limited.
Introduction
In conjunction with genetics, variations in human behaviors such as diet, hand-washing, and vaccination result in differential susceptibility to a range of diseases. Improvements in sanitation and healthcare practices have reduced the mortality and morbidity of many infectious diseases [1]. For example, the implementation of childhood vaccination policies has led to the eradication of smallpox and the elimination of poliomyelitis (polio) in the United States [2–4], and the high efficacy of the measles vaccine, combined with wide vaccine acceptance in developed countries, had resulted in measles previously being targeted for elimination by 2020 [5]. Recent infectious disease trends, however, have illustrated how easily efforts to eradicate diseases can be undermined: over the past decade, there has been a resurgence of vaccine-preventable diseases (VPDs) in developed countries despite the safety and efficacy of vaccines and high overall childhood vaccination rates [6–9]. Outbreaks of VPDs such as measles, pertussis, and polio have been linked to pockets of under-vaccinated or unvaccinated individuals, which often go relatively unnoticed by the public due to high national estimates of vaccination coverage and the assumption of associated herd immunity [8].
Human decisions regarding health-related behaviors, including preventative care, are influenced by both internal and external motivations. The term “local vaccination cultures” describes the shared beliefs among individuals within a community or region about disease etiology, prevention, and treatment, as well as the experiences with health services and vaccination settings that influence individual vaccine decisions [10]. In areas where there exists a pro-vaccination culture, there may nonetheless be individuals who are vaccine hesitant (as defined by [11]), delaying or refusing to use vaccines despite their availability. Vaccine hesitancy is a complex and context-specific individual behavior influenced by factors that shape vaccination culture such as complacency (the belief that vaccination is unnecessary when the perceived risk of VPDs is low), convenience (the accessibility and affordability of vaccines), and confidence (the level of trust in the efficacy and safety of the vaccine, and in the healthcare system) [9,11].
Vaccine hesitancy was named one of the World Health Organization’s ten threats to global health in 2019 [12], as it is believed to be responsible for decreasing vaccination coverage and thus increasing the risk of vaccine-preventable disease outbreaks worldwide.
Vaccine hesitancy and vaccination opposition are, however, not recent phenomena: since the creation of vaccines in the 18th century, public opposition to vaccinations has been based in theology, politics, law, and general skepticism [13–16]. Two persistent themes of vaccine hesitancy include the idea that vaccinations are more harmful than the diseases they intend to prevent (usually occurring at the introduction of new vaccines), and the idea that vaccines may not be necessary during a decline in disease (usually after compulsory vaccination mandates) [13]. With the introduction of compulsory vaccination in the 19th century came the precursors of contemporary vaccination exemptions [17,18]. The Anti-Vaccination League, founded in mid-19th- century London, argued that compulsory vaccination invaded the people’s liberties [19]. A 1969 review of mandatory vaccination in the United States revealed a similar objection—“infringement on personal liberty”—along with an aversion to government intrusion on religious beliefs and a general distrust of medical science [20]. Thus, since the advent of vaccinations, cultural context has interacted with vaccine-related beliefs and in turn influenced vaccination behaviors, ultimately affecting population-level immunity and public health.
Due to developments in media and communication, including widespread engagement with social media platforms, all types of information are more accessible and faster spreading than ever before. Unfortunately, negative information (such as anecdotes about adverse drug reactions), misinformation (such as the discredited link between vaccines and autism), and maladaptive practices (such as delayed vaccination schedules [21,22]) can spread more quickly through these digital channels than by traditional word-of-mouth [23], resulting in the increased adoption of harmful beliefs and practices. In particular, a small but very vocal community, sometimes dubbed “Anti-Vaxxers,” stoke the fears and emotions that contribute to vaccine hesitancy, and their reach is aided by social media [9,24]. Anti-vaccine sentiments are still on the rise despite numerous studies countering, discrediting, and debunking the spurious connection between vaccines and autism [25] and other anti-vaccination arguments [26]. Additionally, individuals are typically more trusting of those with whom they have a personal connection and more skeptical of health providers and scientists [27], therefore anti-vaccine attitudes from one’s social network are more likely to be internalized than pro-vaccine information from healthcare professionals.
Health-related behaviors are also affected by homophily—the tendency of individuals to choose social contacts and mates who are similar to themselves [28–31]. For example, individuals with online social contacts who share similar demographic characteristics had significantly higher adoption of new health-related behaviors introduced in that online environment [32]. In addition, network-based simulations suggest that individuals with similar vaccine-hesitant opinions form groups that are more susceptible to vaccine-preventable diseases, impeding the attainment of herd immunity and substantially increasing the likelihood of disease outbreak in these clusters [33]. Some of the predictions of these simulations have been observed in the real world: for example, a number of recent VPD outbreaks have been linked to tight-knit cultural groups, such as the Ohio Amish community, which has specific teachings on health practices [34,35], and the Minnesota Somali immigrant community, which was vulnerable to anti-vaccination rhetoric [36,37]. If individuals have a tendency toward homophily on the basis of vaccine-related beliefs, preferentially partnering with others who have similar beliefs and mindsets, children might be increasingly likely to have two vaccine-hesitant or two vaccine-confident parents and to inherit their beliefs. When vaccine-related beliefs are clustered on a social network, homophily could further expand and reinforce these clusters as vaccine-hesitant individuals seek out interactions with one another, exacerbating the risk of disease outbreak. Thus, a particular health culture—collective beliefs and behaviors regarding health—could be seen as a cultural “niche” in the sense that it influences an individual’s beliefs, behaviors, and social network connections.
Niche construction is a process in which organisms modify their local environment, thus altering selection pressures on themselves and the other organisms that share that environment [38,39]. In cultural niche construction, humans modify their cultural environments—for example, their beliefs, behaviors, preferences, and social contacts—in ways that subsequently alter evolutionary pressures on themselves and/or their culture [39]. Within various human-constructed cultural niches, for example in agricultural niches, there exist niche-specific differential effects on health and health-related behaviors (for example, the link between dairy farming and lactose tolerance in adulthood [40]. Mathematical models of niche construction have been used to help understand biological evolution, and this type of model has been recently expanded to explain the evolution of cultural behaviors, with applications to religion, fertility, and the evolution of large-scale human conflict [29,30,39,41–43]. The cultural niche framing of health cultures allows us to apply these modeling frameworks to the evolution of human disease response. Modeling the belief-behavior interactions underlying vaccination coverage allows us to better understand how vaccination cultures are formed and how they can be transformed to promote public health.
Compartmental models, such as the Susceptible-Infected-Recovered (SIR) model and its various extensions, are the most established set of epidemiological models used to predict the spread of infectious disease. Even though more epidemiological models have begun to incorporate aspects of human behavior (e.g [44,45]), these models do not typically incorporate the effects of population beliefs and changing cultural landscapes on disease transmission. For example, they do not consider social preferences or early-life influences on adult behaviors, and how these affect the health beliefs that drive specific behaviors. SIR models with a vaccination component (e.g. [46,47]) demonstrate that the introduction of vaccination decreases the susceptible and infected populations proportional to vaccination rate [46]. Though these models are very useful in determining the intensity of intervention needed to address an epidemic [46], they do not explain fluctuations in vaccination rates or lower-than-expected rates of adoption based on cultural factors. In wealthier countries, belief systems act as the main barrier to vaccination, as opposed to lack of vaccine access [33,48]. The perception of vaccine importance and disease risk is a driving factor in vaccine behavior, even for vaccinated individuals [27,49–53]. The rates of non-medical exemption from vaccines (exemption on the basis of religious, philosophical, and personal beliefs), have been increasing in the United States, with some believing the “negative consequences of exemption are not sufficient to justify violating parental autonomy” [54,55]. Thus, understanding and incorporating the underlying health cultures and their evolution, including the interplay between beliefs and behaviors, will allow us to build more comprehensive and representative models of vaccinations dynamics and better support public health efforts.
In this study, we model the development of vaccine hesitancy through a cultural evolution framework, incorporating the transmission of vaccine culture both from parents and from the community. We shed light on the situations in which vaccine hesitancy is most likely to spread and potentially lead to an increase in vaccine-preventable diseases. By modeling the evolution of vaccination behaviors through the lens of cultural niche construction, we aim to assess the dynamic interactions between vaccine beliefs (shaped by social interactions) and behaviors (influenced by these beliefs). With this framework, we can also model the effects of vaccine mandates and of vaccine accessibility on both beliefs and behaviors. In addition, we consider that the perception of the relative risks of a disease and its preventive vaccine can fluctuate based on the prevalence of vaccination, such that the population’s vaccination coverage can influence individual decision-making. We further incorporate homophily, here represented as the likelihood that people with similar vaccine-related beliefs will preferentially mate with one another, to understand how social subcultures influence behaviors.
Beliefs influence but do not dictate an individual’s behaviors, and these belief-behavior interactions are influenced by the broader cultural landscape. We propose that scientific research can better address how to mitigate VPD outbreaks by understanding the cultural dynamics of vaccine hesitancy. In this manuscript, we aim to take a cultural approach to understanding the evolution of vaccine hesitancy and its interactions with vaccination coverage and vaccine-preventable disease using a generalizable modeling framework for belief-behavior interactions.
Methods
To model the cultural niche construction of vaccine beliefs and behaviors, we build on the framework of [30] to assess the effects of vaccine attitudes on vaccination behaviors and on the resulting vaccination culture. We explore how vaccination patterns evolve in a population when a cultural trait, such as vaccine hesitancy, can influence but not perfectly predict a behavior, such as vaccinating one’s children.
We consider two cultural traits: V, a vaccination trait, and A, a vaccine attitude trait. Each trait has two possible states, V+ or V− and A+ or A−, respectively. V+ individuals are vaccinated and V− individuals are unvaccinated, and A+ individuals are vaccine confident and A− individuals are vaccine hesitant; thus, there are four possible phenotypes: V+A+ (type 1: vaccinated and confident), V+A− (type 2: vaccinated and hesitant), V−A+ (type 3: unvaccinated and confident), and V−A− (type 4: unvaccinated and hesitant), whose population frequencies are denoted by x1, x2, x3, and x4, respectively, with 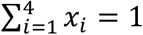 .
.
The attitude trait (A) can influence the dynamics of the vaccination trait (V) in two ways: by affecting the likelihood that couples vaccinate their offspring, and by determining with whom each adult will preferentially pair in assortative interactions. The state of the vaccine attitude trait (A) informs the value of an assortative mating parameter (αk), which measures the departure from random mating. We define a ‘choosing parent,’ arbitrarily assigned as the first member of each mating pair. The choosing parent’s A state dictates the level of assortative mating, that is, the degree to which an individual of a given A state will preferentially mate with another individual of the same state, expressed by parameters αk where k = {1, 2} and 0≤αk≤1 (Table S1). If the choosing parent is A+, this individual mates preferentially with other A+ individuals with probability α1, and mates randomly with probability 1−α1, whereas if the choosing parent is A−, this individual mates preferentially with other A− individuals with probability α2, and mates randomly with probability 1−α2. There are sixteen possible mating pairs from the four phenotypes described, and we use the notation mi,j to indicate the frequency of a mating between a choosing parent of type i and the second parent of type j where i, j = {1, 2, 3, 4} (Table S1); in other words, m1,3 represents the mating frequency of V+A+ (x1) and V−A+ (x3).
Since the two traits in question (A and V) are transmitted vertically, for each phenotype we must specify the probability that the mating produces an offspring of that phenotype. The vaccine confidence trait (A+) is transmitted with probability Cn, and the vaccine hesitancy trait (A−) is transmitted with probability 1−Cn (for n = {0, 1, 2, 3} as shown in Tables 2 and S2). If C0 = 0, two A− parents will always produce A− offspring, and if C3 = 1, two A+ parents will always produce A+ offspring. However, if C0 > 0, two A− parents can produce A+ offspring at some probability, and similarly if C3 < 1, two A+ parents can produce A− offspring with some probability.
Transmission of vaccination (V+ with probability Bm,n for m, n = {0, 1, 2, 3}; Table 1) is more complex, since parents’ vaccine attitudes (A), in addition to their own vaccination states (V), can influence their behavior in vaccinating their offspring via a set of “influence parameters” that inform vaccination probabilities. The probability that each mating pair produces an offspring with the V+ trait (i.e. vaccinates their offspring) is a scaled product of the influence of parental attitudes (cn for n = {0, 1, 2, 3}) and the influence of parental vaccination states (bm for m = {0, 1, 2, 3}) (Tables 2 and S2). For example, for mating pair V+A+ × V+A−, their combined vaccination states (V+ × V+) will influence vaccination behavior by b3, and their combined attitude states, (A+ × A−), will influence vaccination behavior by c2. Therefore, a V+A+ × V+A− mating will produce a V+ offspring with probability 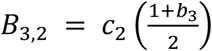 ; this pair will also produce an A+ offspring with probability C2 based on their combined attitude states. Thus, according to the model, this pairing will produce a V+A+ offspring with probability B3,2C2 and a V+A− offspring with probability B3,2(1−C2). We note that assortative mating (αk>0) will increase the frequency of matings between individuals that share an attitude trait, with these non-random interactions in turn skewing vaccination outcomes in line with those of same-state couples (via c0 and c3).
; this pair will also produce an A+ offspring with probability C2 based on their combined attitude states. Thus, according to the model, this pairing will produce a V+A+ offspring with probability B3,2C2 and a V+A− offspring with probability B3,2(1−C2). We note that assortative mating (αk>0) will increase the frequency of matings between individuals that share an attitude trait, with these non-random interactions in turn skewing vaccination outcomes in line with those of same-state couples (via c0 and c3).
Demonstrating the trait presence (+) and absence (–) combinations associated with m, n subscripts. For example, the + × – combinations is associated with m and n subscript value 2: an A+ × A− pairing transmits A+ at probability C2. This rule applies to parameters Cn, bm, Bm,n, cn, as shown in Table S2.
Transmission and influence probabilities are constant throughout a single simulation, with values ranging from 0 to 1. At default settings, the influence parameters bm and cn, and the transmission parameter Cn would take the following values: C0, b0, c0 = 0.01; C1, C2, b1, b2, c1, c2 = 0.5; and C3, b3, c3 = 0.99. In our model, parental vaccine beliefs (cn) have a greater influence than their own vaccination status (bn) on their likelihood of vaccinating their offspring, so offspring vaccination is guaranteed at some probability only if cn > 0.
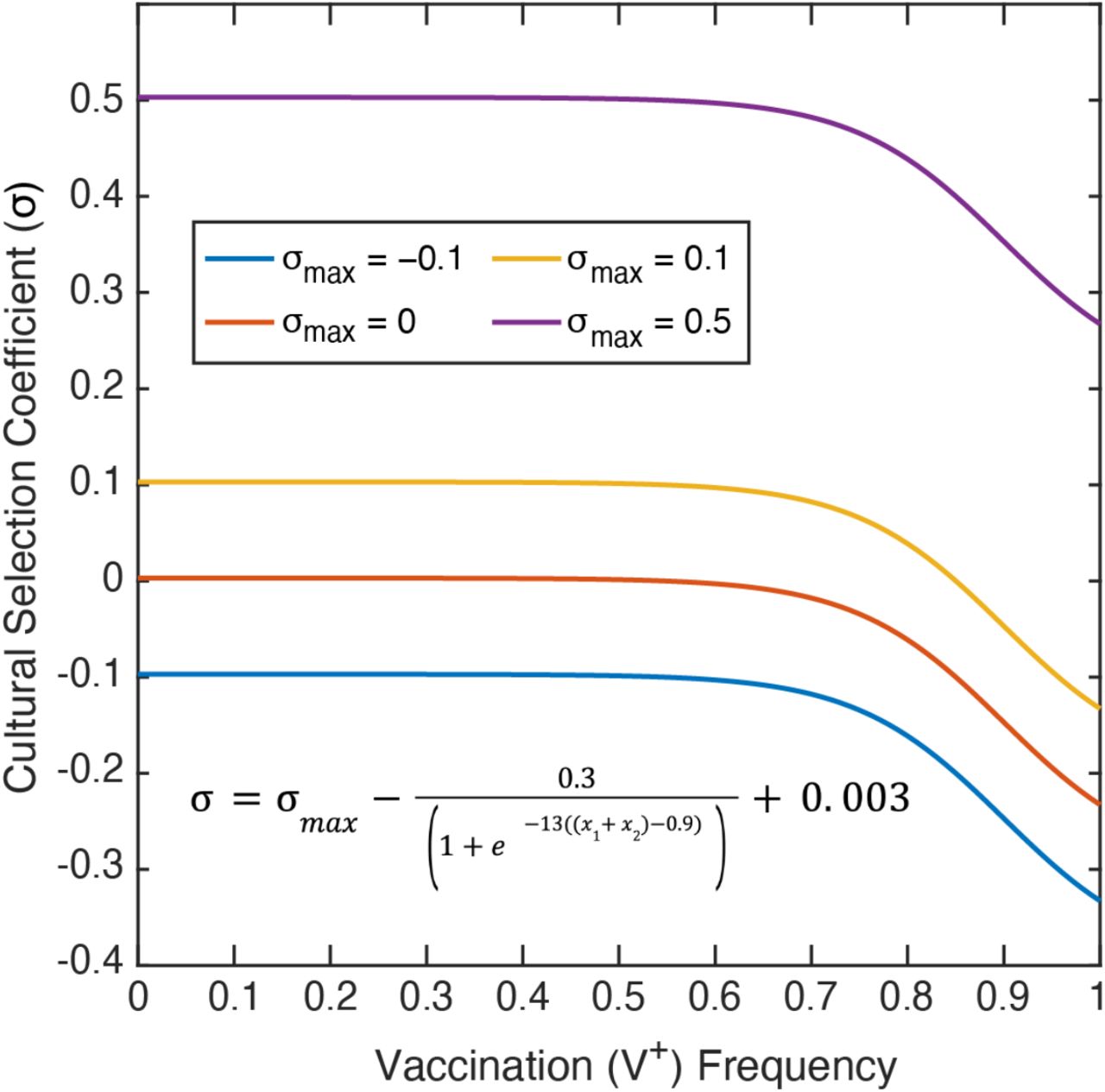
The cultural selection pressure on vaccination is given by the parameter σ, such that the frequency of the V+A+ and V+A− phenotypes are multiplied by 1+σ after vertical cultural transmission has occurred. At the end of each timestep, the frequency of each phenotype is divided by the sum of all four frequencies, ensuring that the frequencies sum to 1. This cultural selection coefficient is implemented in the same way as a selection coefficient in a population-genetic model, but unlike the latter, it is structured to encompass both biological fitness and cultural selection pressures, including perceived risks or benefits of the vaccine itself, personal cost-benefit analyses of preventative health behaviors, and the structural or societal-level factors influencing vaccination rates [56,57]. Since the frequencies of V+ phenotypes are multiplied by 1+σ, this parameter modulates whether there are more or fewer vaccinated individuals than expected: in other words, when σ>0, vaccinated individuals are more common in a set of offspring than would be expected strictly by parental beliefs and vaccination statuses. We calculate σ in each timestep as a function of the current vaccination coverage (frequency of V+, i.e. x1 + x2), and in each simulation we specify σmax as the maximum cultural selection pressure of getting vaccinated (−1≤σmax≤1) (see the cultural selection coefficient function in Figure 2). This function was constructed by fitting a curve to pre-specified conditions: we assume that when vaccination coverage is low, the real and perceived benefits of vaccination are highest and thus the cultural selection pressure is near σmax, however, as vaccination coverage increases, the perceived benefits of vaccination decrease and the cultural selection pressure is reduced (Figure 2).
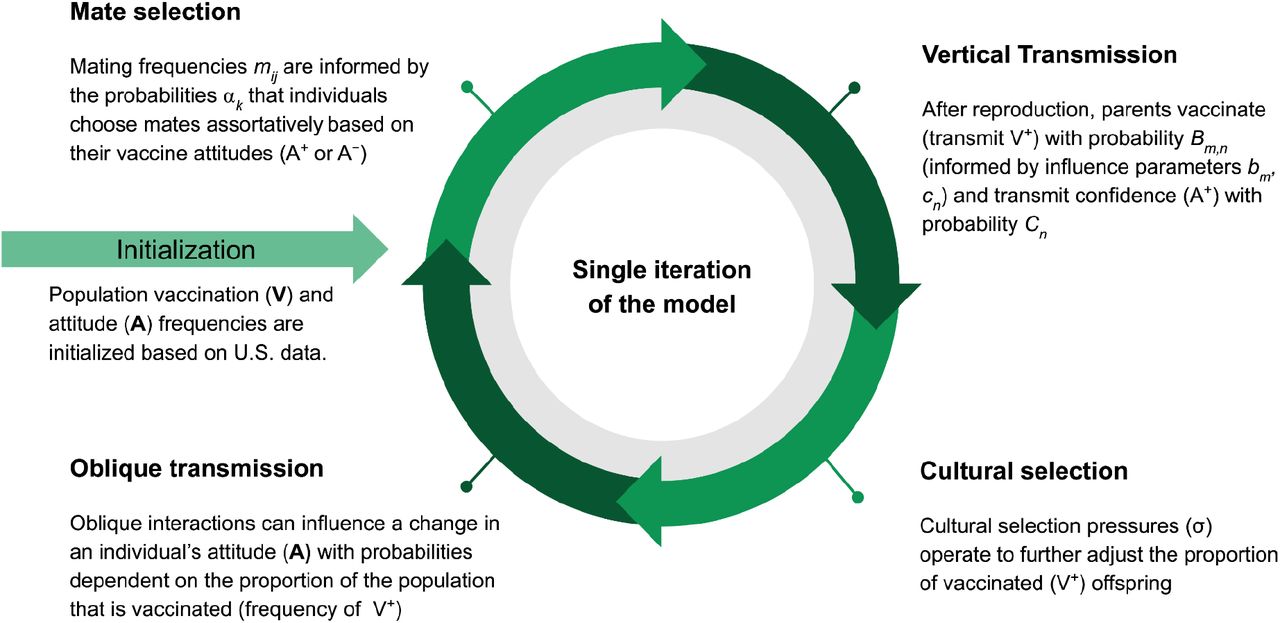
The schematic shows the processes within a single model iteration. The model is initialized with the phenotypic frequencies (V+A+, V+A−, V−A+, V−A−) in the population. After mate selection, individuals reproduce; they then vertically transmit vaccination and attitude traits to their offspring. Vaccination trait frequencies are further modulated by cultural selection. Oblique transmission (cultural transmission from non-parental adults in the population) follows, in which offspring may alter their attitude state. (Parameters, their definitions, and default values are listed in Table 1.)
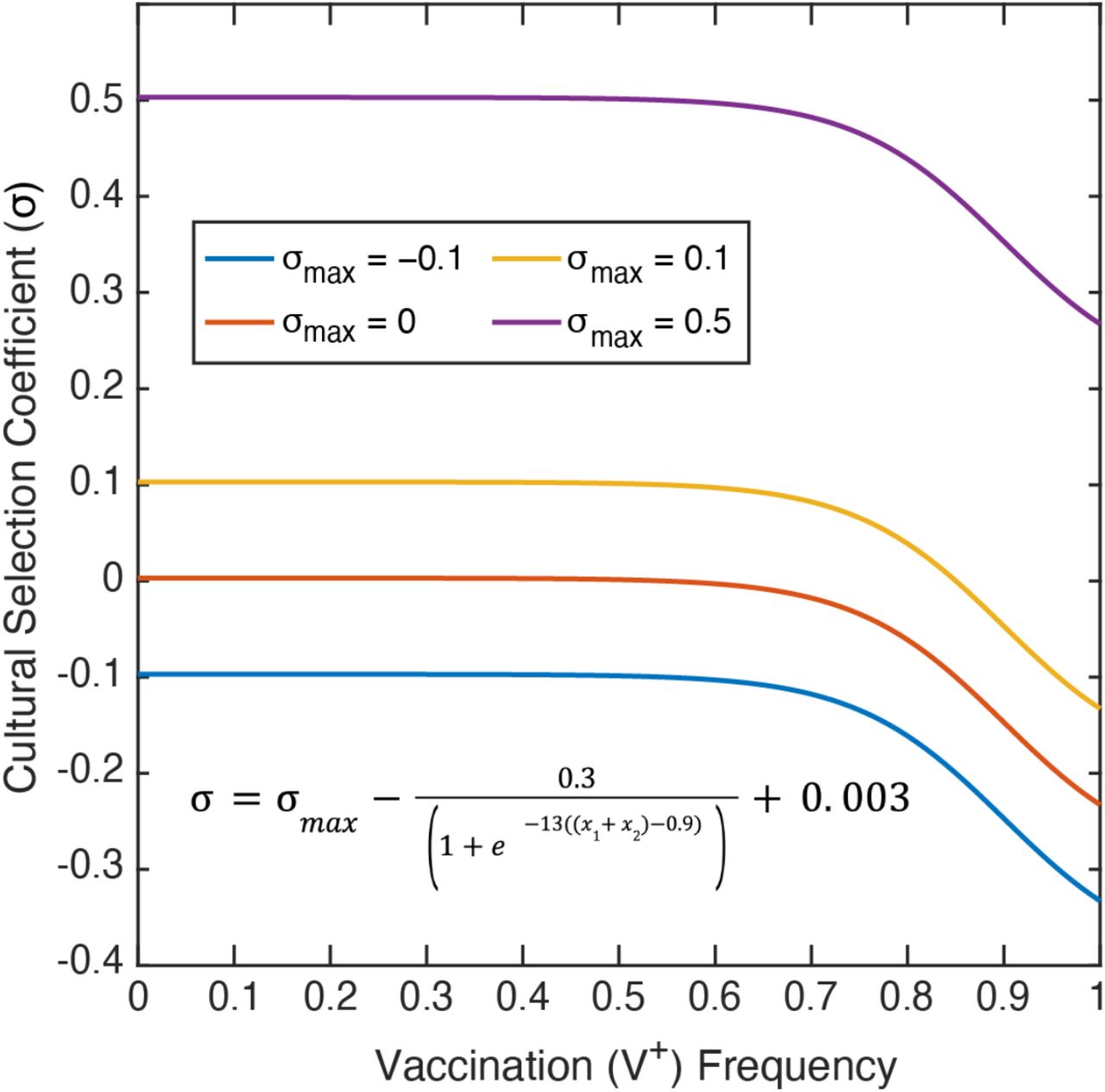
The cultural selection coefficient function was constructed by fitting a curve to specified conditions, and considers both health and non-health related effects. The selection coefficient (σ; vertical-axis) is dependent on the frequency of vaccinated individuals (V+) in the population (horizontal-axis). σmax is the maximum cultural selection coefficient associated with being vaccinated. Perceived vaccine benefit is reduced as vaccination coverage increases, since the negative effects of the disease will be less apparent. Note: Of the σmax values shown, only σmax = 0.1 allows the cultural selection pressure to be either positive or negative at a given timepoint depending on the frequency of vaccination.
Thus far, we have described vertical cultural transmission from parent to offspring. The model incorporates a second phase with oblique cultural transmission (i.e. influence from non-parental adults), in which individuals can change their inherited vaccine attitudes (A) due to influence from other adults in the population. There are two probabilities associated with attitude modulation: the probability that an vaccine hesitant (A−) individual adopts the vaccine confident (A+) state (A− to A+ transition probability, given by A→Confident in Figure 3), and the probability that an A+ individual adopts the A− state (A+ to A− transition probability, given by A→Hesitant in Figure 3). As with the strength of cultural selection (σ) described previously, the probability that offspring change their vaccine attitude is a function of the V+ frequency in the population. As the frequency of vaccinated individuals (V+) increases in the population, vaccine-confident individuals (A+) are more likely to become hesitant (A→Hesitant probability increases) and vaccine-hesitant individuals (A−) are less likely to become confident (A→ Confident probability decreases). By modulating the attitude transition probabilities according to the vaccination coverage in this manner, we assume that when vaccine coverage (V+ frequency, x1 + x2) is low, disease occurrence is high and the negative effects of the disease are experienced widely, thus the benefits of being vaccinated (and the costs of not being vaccinated) are more evident [58,59]. As vaccination coverage (V+) increases in the population, and thus disease occurrence is low, the benefits to being vaccinated are less obvious, while low-probability costs such as adverse reactions become more apparent and could be perceived as being riskier than the disease itself. Modulating both the attitude transition probabilities and the cultural selection coefficient according to the level of vaccination coverage in a population reflects that perceptions about the vaccine and its associated effects on health could be meaningfully different in a population with high vaccination coverage than in one with low coverage.

Attitude transition probability functions were constructed by fitting a curve to specified values. Attitude transition probability (vertical axis) is a function of the vaccination frequency in the population (V+; horizontal axis). The probability that a vaccine hesitant individual adopts vaccine confidence (A− to A+ transition probability, shown in black) is determined by the function A→Confident, and the probability that a vaccine confident individual adopts vaccine hesitancy (A+ to A− transition probability, shown with a blue dashed line) is determined by the function A→Hesitant.
To compute the frequency of a given phenotype in the next iteration, we sum the probability that each mating pair produces offspring of that phenotype over each of the sixteen possible mating pairs. Cultural selection (σ), described above, then operates on offspring with the V+ trait. The full recursions, giving xi′ phenotype frequencies in the next iteration in terms of xi in the current iteration, are given in Text S1. If xi′is equal to xi, the system is at equilibrium. Unless otherwise stated, the model is initialized with phenotypic frequencies structured to represent those of the United States: x1 (frequency of V+A+) = 0.81, x2 (V+A−) = 0.1, x3 (V−A+) = 0.07, x4 (V−A−) = 0.02. These frequencies were estimated using reports of Measles-Mumps-Rubella (MMR) vaccination rates and estimates of vaccine attitude frequencies obtained from various sources in the literature [60,61] and the Centers of Disease Control ChildVax database [62,63].
Results
Parent-to-Offspring Interactions (Simulations with vertical transmission only)
We began by testing this model with only vertical transmission dynamics, i.e. only parent-to-offspring transmission, varying parameter values in turn to test their effects on population vaccination behavior and attitudes. In the vertical transmission phase of the model, parents choose whether to vaccinate their offspring (i.e., transmit V+) or to not vaccinate (V−), and parents also transmit a vaccine attitude (confidence, A+, or hesitancy, A−), each with a specified probability given the phenotypes of the parents. The parental attitude state, vaccination status, assortative mating levels, and cultural selection parameters interact to affect vaccination coverage (frequency of V+) and vaccine confidence (frequency of A+). Couples with mixed vaccination and/or attitude states (V+ × V−, A+ × A−) are assumed to be more variable in their decision to vaccinate their offspring than parents who share the same state. Hence, we primarily modulated probabilities associated with these mixed-phenotype pairings for the purposes of testing our model.
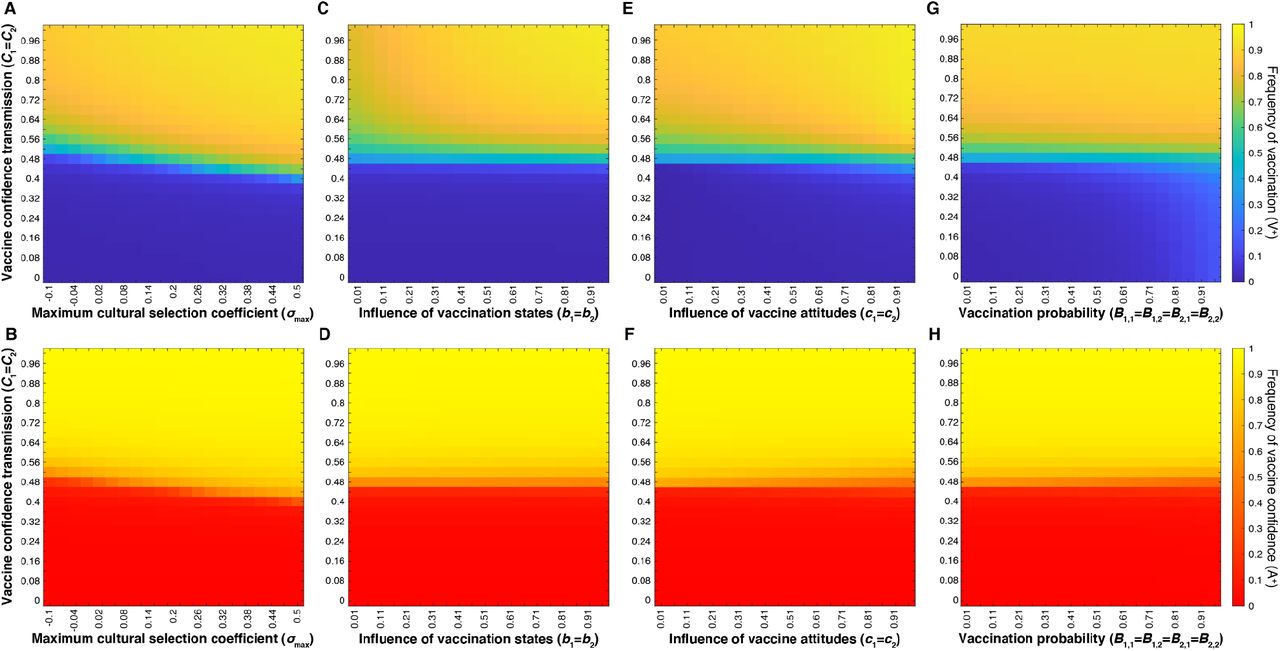
First, we compared the effects of varying the confidence transmission probabilities for mixed-attitude couples (C1 and C2), i.e. those with one vaccine-hesitant individual and one vaccine-confident individual, in combination with multiple factors: 1) the maximum cultural selection coefficient (σmax) (Figure 4A-B), 2) the vaccination influence parameters b1 and b2 (Figure 4C-D), 3) the attitude influence parameters c1 and c2 (Figure 4E-F), and 4) the vaccination probabilities of couples with mixed traits, B1,1, B1,2, B2,1, B2,2 (Figure 4G-H). In each examination, we observed a Cn threshold: there is a mid-range value of Cn at which vaccination coverage and vaccine confidence traits are polymorphic (i.e. both forms of each trait coexist in the population), separating definitive high (⪆80%) and low (⪅30%) levels of vaccination coverage and confidence. This Cn threshold value is more sensitive to σmax than to bm, cn, or Bm,n: the threshold value is lowered as σmax increases (diagonal line in Figure 4A-B). Although vaccination probability (Bm,n) is dependent on both cn, the influence of parental vaccine attitude, and bm, the influence of parental vaccination state (Table S2), modulating either type of influence of mixed-state parents has little effect on the level of vaccination coverage and negligible effects on confidence levels at each non-threshold Cn (Figure 4C-F).

Heatmaps showing final vaccination coverage and vaccine confidence after 100 time-steps with no oblique transmission, only parent-to-offspring transmission. The top row (A, C, E, G) shows vaccination coverage (i.e. frequency of V+ in the population) with low coverage in blue and high coverage in yellow; the bottom row (B, D, F, H)) shows the corresponding final vaccine confidence (i.e. frequency of A+), with low confidence in red and high confidence in yellow. Unless varied on the horizontal or vertical axis, other parameters are set to the default values given in Table 1. In our model, parents’ likelihood of vaccinating their children depends on both their vaccination state and their attitude state. This figure shows that the strength of parental transmission of vaccine confidence (Cn) has a much stronger effect on the equilibrium levels of both vaccine coverage (V+) and confidence (A+) than other parameters: the maximum cultural selection coefficient, σmax (A,B), the influence of parental vaccination state, bm (C, D), the level of influence of parental vaccine attitudes on their vaccination behaviors, cn (E,F), and the probability that mixed-state parents vaccinate their offspring Bm,n (G,H).
Interestingly, direct modulation of the mixed-trait couple vaccination probability (B1,1 = B1,2 = B2,1 = B2,2) also has little power in affecting coverage and confidence levels at equilibrium (Figure 4G-H). We hypothesize that predominantly high or predominantly low confidence transmission within a population reduces the occurrence of “mixed-trait” pairings, i.e. if the majority of the population becomes confident or hesitant, there are fewer confident-hesitant and vaccinated-unvaccinated pairings. Thus, the effect of modulating mixed-trait vaccination probabilities (B1,1, B1,2, B2,1, B2,2) is significantly minimized as these couples approach low frequencies in the population, and confidence transmission dominates the vaccination patterns. To further explore this relationship, we varied all vaccination probabilities (Bm,n), not just those of mixed couples, while modulating mixed trait confidence transmission probabilities (C1 = C2) as before. To directly alter vaccination probabilities while still accounting for the couple’s vaccine attitudes, we set ranges of values for Bm,n that vary along the horizontal axis of Figure 5, with the vaccination probability for two hesitant parents (e.g. B0,0) on the lower end of the range and the vaccination probability for two confident parents (e.g. B3,3) on the higher end of the range (Table S3). In these tests, we observe increasing equilibrium vaccination coverage as Bm,n probabilities increase, with higher coverage in high-confidence transmission environments (Figure 5). If we vary both confidence transmission parameters and vaccination probability parameters by implementing range shifts in both Cn and Bm,n, we observe an interaction between confidence transmission and vaccination probability that determines vaccination coverage (Figure S1). In both, we confirm vaccination coverage levels are determined by an interaction between confidence transmission and vaccination probability, whereas confidence levels are dictated primarily by levels of confidence transmission. In sum, the degree to which parents with mixed vaccine-hesitant and vaccine-confident attitudes transmit vaccine confidence instead of vaccine hesitancy to their offspring is a key factor in determining population trait majorities which can drastically shift population dynamics.

Heatmaps showing final vaccination coverage (A) and corresponding vaccine confidence (B) after 100 time-steps with no oblique transmission. Confidence transmission in mixed-attitude couples (C1 = C2) is varied along the vertical axis, while the vaccination probabilities (Bm,n) are set within the range indicated on the horizontal axis, with B0,0, B1,0, B2,0 and B3,0 taking the lowest value and B3,3 taking the highest value (Table S3). We show increased equilibrium vaccination coverage with increasing vaccination probability ranges, while confidence levels are primarily dictated by proportion of the population transmitting confidence or hesitancy.
Next, we hold vaccine confidence transmission (Cn) at default probabilities, reminiscent of Mendelian transmission, such that two vaccine confident or two vaccine hesitant parents predictably transmit their vaccine attitude, and parents with differing vaccine attitudes each have a ∼50% chance of transmitting their phenotype, e.g. C0 near 0, C1 and C2 at 0.5, C3 near 1 (Table 1). We then varied cultural selection in combination with vaccination-associated probabilities (bm, cn, Bm,n). With Cn held constant, cultural selection (σmax) is the primary factor determining vaccination coverage and confidence levels (Figure 6. Raising the maximum cultural selection coefficient increases the equilibrium level of vaccination coverage and vaccine confidence across various levels of vaccination state influence (bm) (Figure 6A-B), vaccination attitude influence (cn) (Figure 6C-D), and vaccination probability (Bm,n) (Figure 6E-F). Unlike in Figure 4, vaccine confidence does not always mirror vaccination coverage across all levels of attitude influence (cn) or vaccination probabilities. Instead, vaccine confidence levels decline with increased cn and increased Bm,n for σmax ⪅ 0.3 (Figure 6D, F), as well as for both increased cn and increased bm (Figure S2. This dynamic is interesting as these parameters influence vaccination behavior, hinting that high vaccination rates could reduce a populations’ expected vaccine confidence. Vaccination coverage and vaccine confidence remain low when cultural selection does not favor vaccination (σmax ⪅ 0), i.e. parents vaccinate their children at or below the levels expected based on cultural transmission rates.
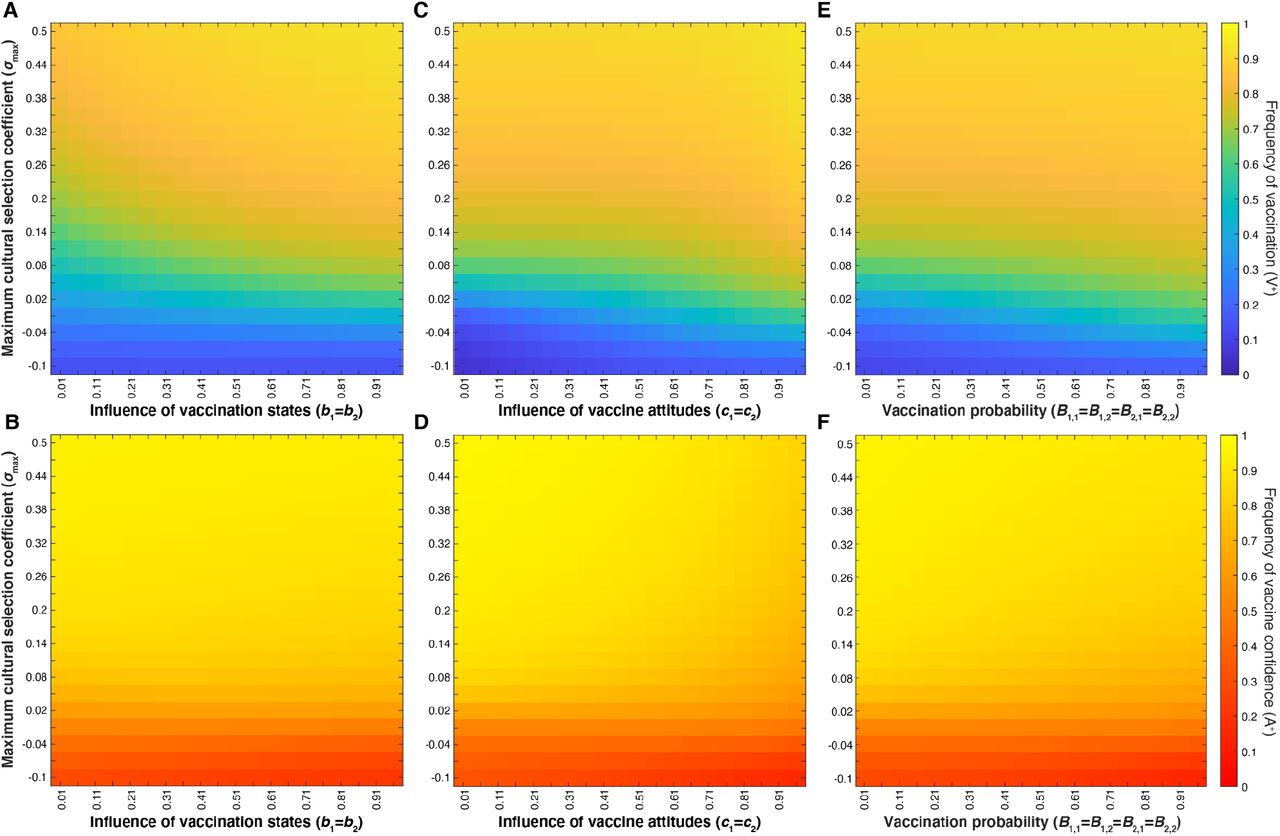
Heatmaps showing final vaccination coverage (A, C, E) and final vaccination confidence (B, D, F) after 100 time-steps with no oblique transmission, only parent-to-offspring transmission. As in Figure 4, parameters not varied here are given in Table 1. Parents’ likelihood of vaccinating their children depends on both their vaccination state and their attitude state. At default probabilities of vaccine confidence transmission (Cn values in Table 1), these figures show that modulating the maximum cultural selection coefficient affects the equilibrium levels of vaccination coverage and vaccine confidence across the range of specified parameters: parental vaccination state influence, bm (A, B), parental attitude state influence, cn (C,D), and offspring vaccination probability, Bm,n (E,F). Unless directly modulated (as in panels E-F), Bm,n varies with bm and cn (Bm,n = cn((1+bm)/2).
Temporal dynamics of vaccine-related beliefs and behaviors
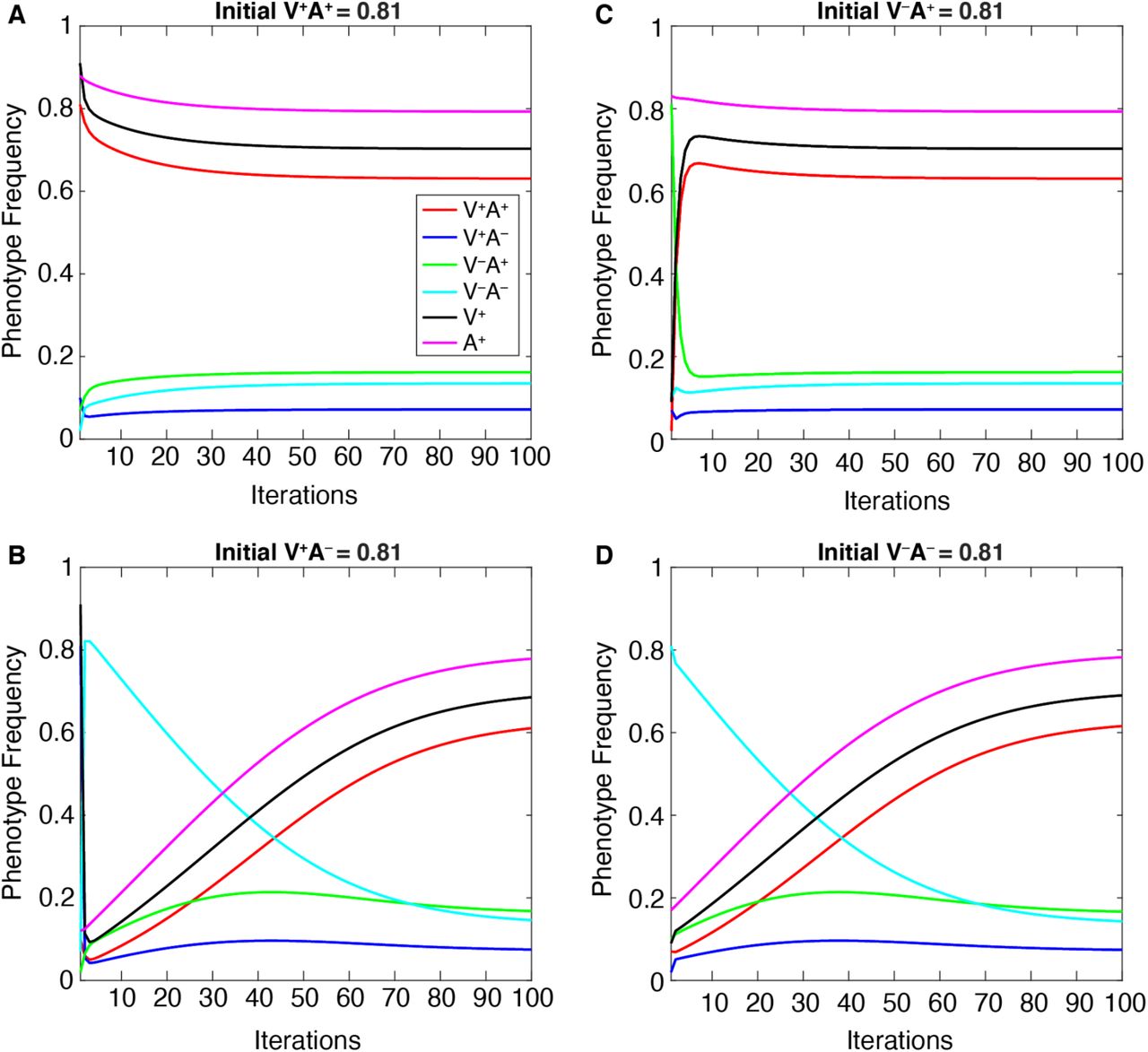
To test whether the equilibrium phenotype frequencies were sensitive to starting frequencies, we plotted the dynamics of each phenotype over time at default parameters (given in Table 1). For each set of initial phenotype proportions tested, each phenotypic frequency quickly adjusted to approach equilibrium values and then gradually plateaued to a stable equilibrium (vertical transmission: Figure 7 and Figure S3, vertical+oblique transmission: Figure S4. This demonstrates that equilibrium frequencies of vaccination coverage and vaccine confidence are determined by the parameter conditions rather than by the initial frequencies themselves.

The change in each of the four phenotype frequencies and the total V+ and A+ frequencies (vertical axis) over 100 iterations of the model (horizontal axis). Initial frequencies are varied, such that we begin each simulation with a different phenotype at an initial high frequency (0.81): V+A+ in panel A, V+A− in panel B, V−A+ in panel C, V−A− in panel D; the remaining phenotypes are set to lower frequencies (0.1, 0.07, 0.02). See Figure S3 for a full listing of these initial frequencies. The remaining parameters are held at default values (Table 1).
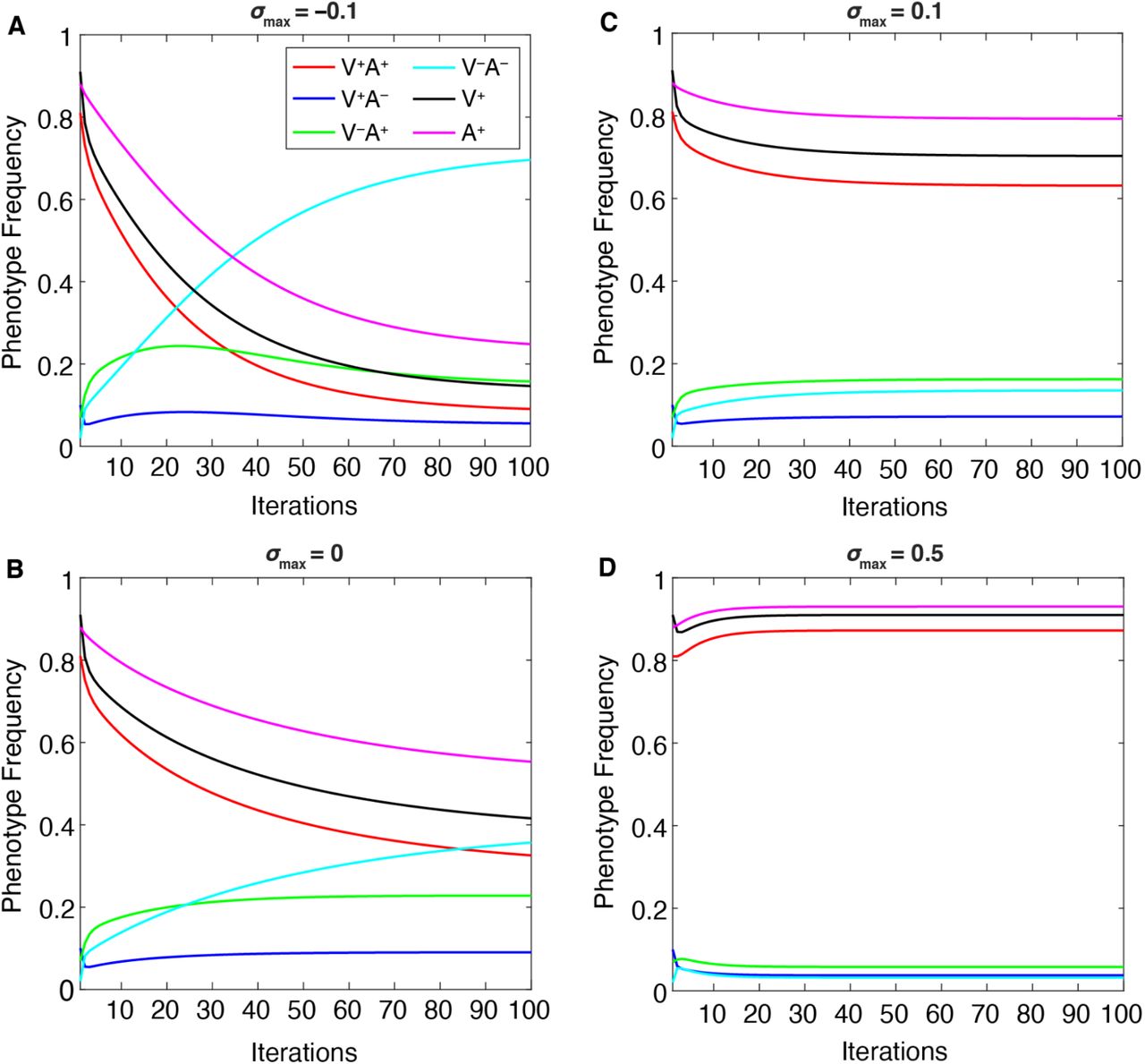
When two parameters in particular are varied—maximum cultural selection (σmax) or confidence transmission (C1 = C2)—we observe a trade-off between the V−A− phenotype, which dominates at lower values of these parameters, and the V+A+ phenotype, which dominates at higher values (Figures 8–9). Interestingly, the “conflicting” phenotypes (when an individual’s attitude toward vaccinating their children does not match their own vaccination state: V−A+ and V+A−) are present at their highest frequencies at neutral cultural selection (σmax = 0, Figure 8B) and/or neutral confidence transmission (C1 = C2 = 0.5, Figure 9B). Vaccinated individuals have the same fitness regardless of their attitude (V+A+ bears the same selection pressure as V+A−), so it is worth noting that at higher levels of confidence transmission and cultural selection, V+A+ increases in frequency but V+A− decreases in frequency (compare Figure 8B-C, Figure 9B-C). This pattern seems to reflect their differing likelihoods of vaccinating their offspring: across all possible partners, vaccinated but vaccine-hesitant parents (V+A−) are less likely to vaccinate their offspring than vaccinated and vaccine-confident parents (V+A+), resulting in more V− offspring. Thus, when V+ is favored by cultural selection, there is indirect selection against the V+A− phenotype (Figure 8). Similarly, indirect selection against V−A+ occurs when V− is favored by cultural selection (Figure 8A): compared to Figure 8B, we observe an increase in V−A− individuals but a decrease in V−A+ individuals, who because of their vaccine confidence have more V+ offspring, which are culturally disfavored in this environment. When cultural transmission from non-parental adults (oblique transmission) was included, described in following sections, we observed similar patterns, but the final equilibria were more likely to be polymorphic, with vaccinated, unvaccinated, confident, and hesitant phenotypes stabilizing at more moderate frequencies than they would have with only vertical transmission (Compare Figures 8–9 to Figures S5-S6).
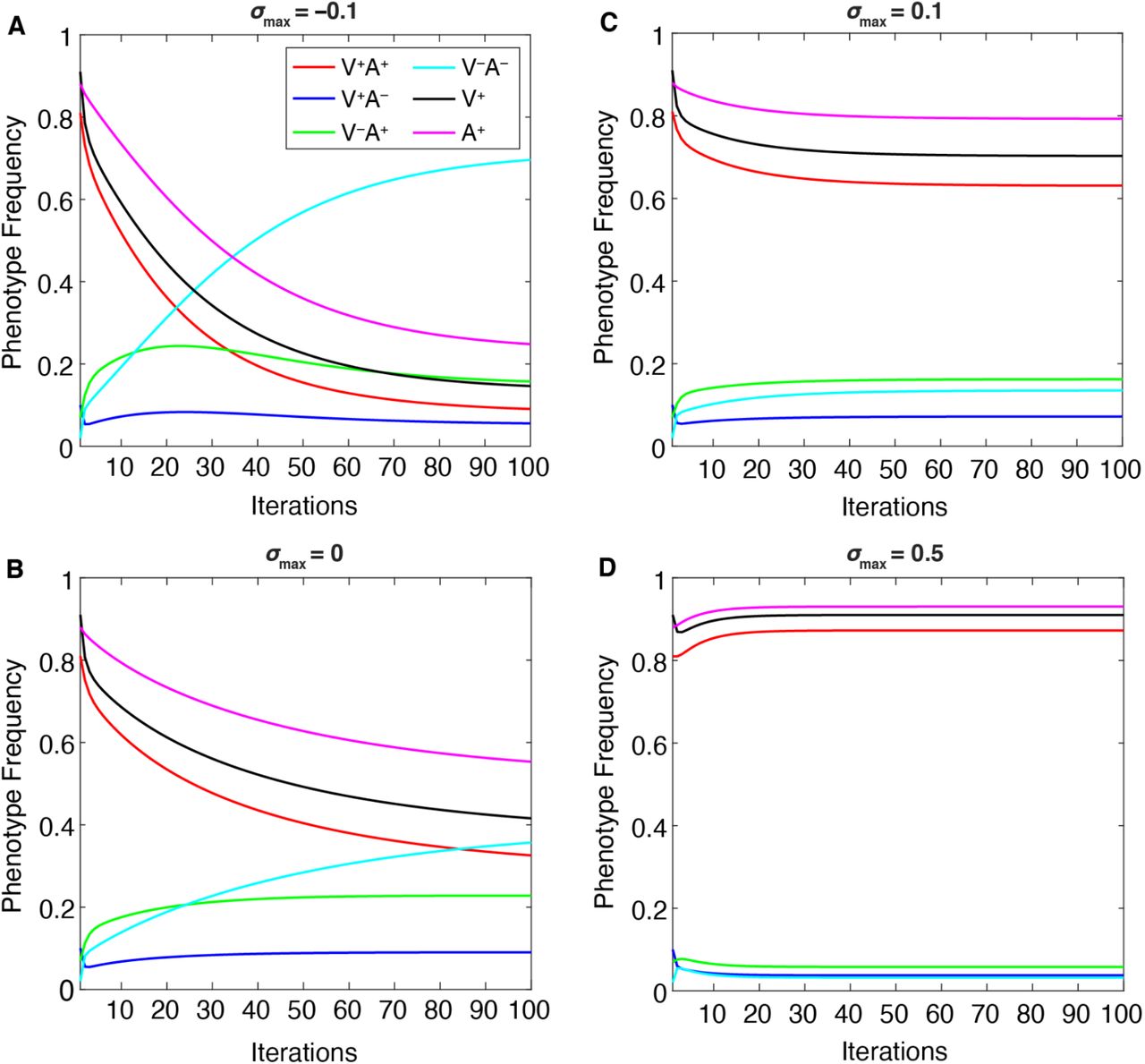
The change in equilibrium phenotype frequencies as the maximum cultural selection coefficient (σmax) is varied: A. σmax = −0.1; B. σmax = 0; C. σmax = 0.1; D. σmax = 0.5, while other parameters are held at default values (Table 1). Cultural selection against vaccinated individuals increases the frequency of V−A−, decreasing the other frequencies (A), whereas increased cultural selection favoring vaccinated individuals increases V+A+ frequencies while decreasing the other frequencies (B, C, D).

The change in phenotype frequencies over 50 iterations as vaccine confidence transmission in mixed couples (C1 = C2) is varied (A. C1 = C2 = 0.1; B. C1 = C2 = 0.5; C. C1 = C2 = 0.8) while other parameters are held at default values (Table 1). The population equilibrates at over 90% A−V− at low confidence transmission (A). Increasing the probability of confidence transmission results in less vaccine hesitancy and, in turn, higher vaccination frequencies (V+A+).
Low confidence transmission (C1 = C2 = 0.1, Figure 9A) increases the frequency of vaccine hesitancy (A−) in the population over time, increasing the probability that more couples choose not to vaccinate their offspring. However, the increase in vaccine hesitancy does not occur equally in vaccinated and unvaccinated individuals: A− frequency may increase overall in this environment, but V+A− frequencies are lower and V−A− frequencies are higher (compared to Figure 9B-C and Figure S6). At neutral confidence transmission probabilities (i.e. when couples with one confident and one hesitant parent are equally likely to transmit either attitude), there is a higher chance that the vaccinated but vaccine-hesitant (V+A−) phenotype is replenished.
However, if confidence is highly transmitted (C1 = C2 = 0.8), the V+A− frequency will be reduced, as this phenotype is more likely to produce A+ offspring than A−, thus increasing V+A+ phenotype frequencies in the population (Figure 9 and Figure S6). If we turn to the other conflicting phenotype, unvaccinated but vaccine-confident (V−A+) individuals become more common when A+ increases in frequency in the population as C1 = C2 increases from 0.1 to 0.5 (Figure 9 and Figure S6). In contrast, higher vaccine confidence transmission (C1 = C2 = 0.8) can lead to a vaccination-promoting environment in which V− frequencies are reduced over time; thus the V−A+ phenotype becomes rare and V+A+ predominates (Figure 9 and Figure S6).
Offspring can Change their Inherited Hesitancy State (Vertical and Oblique Dynamics)
Increased exposure to the attitudes of the broader community (i.e. oblique cultural transmission from non-parental adults in the population) could influence and change vaccination beliefs inherited in childhood. Therefore, we next included these oblique effects in our model to understand how they might modulate vaccine confidence and vaccination coverage levels. In the oblique transmission phase of the model, offspring can change their vaccine attitude with some probability based on the frequency of vaccination in the population (Figure 3). Thus, in addition to the vertical transmission of attitudes and behaviors, phenotype frequencies are further affected by the probability that adult offspring change their attitude (i.e. transition from vaccine confident (A+) to hesitant (A−) and vice versa).
The addition of oblique dynamics produces a pattern of vaccination coverage and vaccine confidence similar to that of simulations run with solely vertical transmission (Figure 4 and Figure 6 compared to Figure 10 and Figure 11, and Figures 7–9 compared to Figures S4-6)—the level of (vertical) vaccine confidence transmission still largely determines the level of vaccination coverage and vaccine confidence (Figure 10). However, oblique cultural influences expanded the polymorphic space, resulting in a wider range of intermediate Cn in which the different phenotypes (vaccinated, unvaccinated, confident, and hesitant) are present in the population in roughly equal proportions. In other words, there is a wider horizontal stripe of moderate values between the definitively high and definitively low equilibrium frequencies in Figure 10 than in Figure 4). Overall, the addition of oblique transmission appears to lead to less polarized results, moving the equilibrium levels of vaccination coverage and vaccine confidence away from extreme values.

Heatmaps showing final vaccination coverage (i.e. frequency of V+ in the population, with low coverage in blue and high coverage in yellow (A, C, E, G)) and final vaccine confidence (i.e. frequency of A+, with low confidence in red and high confidence in yellow (B, D, F, H)) after 100 time-steps in which oblique transmission of vaccine attitude can occur after parent-to-offspring transmission. The likelihood that individuals change their vaccine beliefs depends on the current vaccination coverage of the population (Figure 2). Unless varied on the horizontal or vertical axes, other parameters are set to the default values given in Table 1. Parents’ likelihood of vaccinating their children depends on both their vaccination state and their attitude state; these figures show that the strength of parental transmission of vaccine confidence (Cn) has a much stronger effect on the equilibrium levels of both vaccine coverage (V+) and confidence (A+) than other tested parameters do: the maximum cultural selection coefficient, σmax, (A,B), the influence of parental vaccination state, bm, (C, D), the level of influence of parental attitudes on their vaccination behaviors, cn, (E,F), and offspring vaccination probability, Bm,n (G,H).
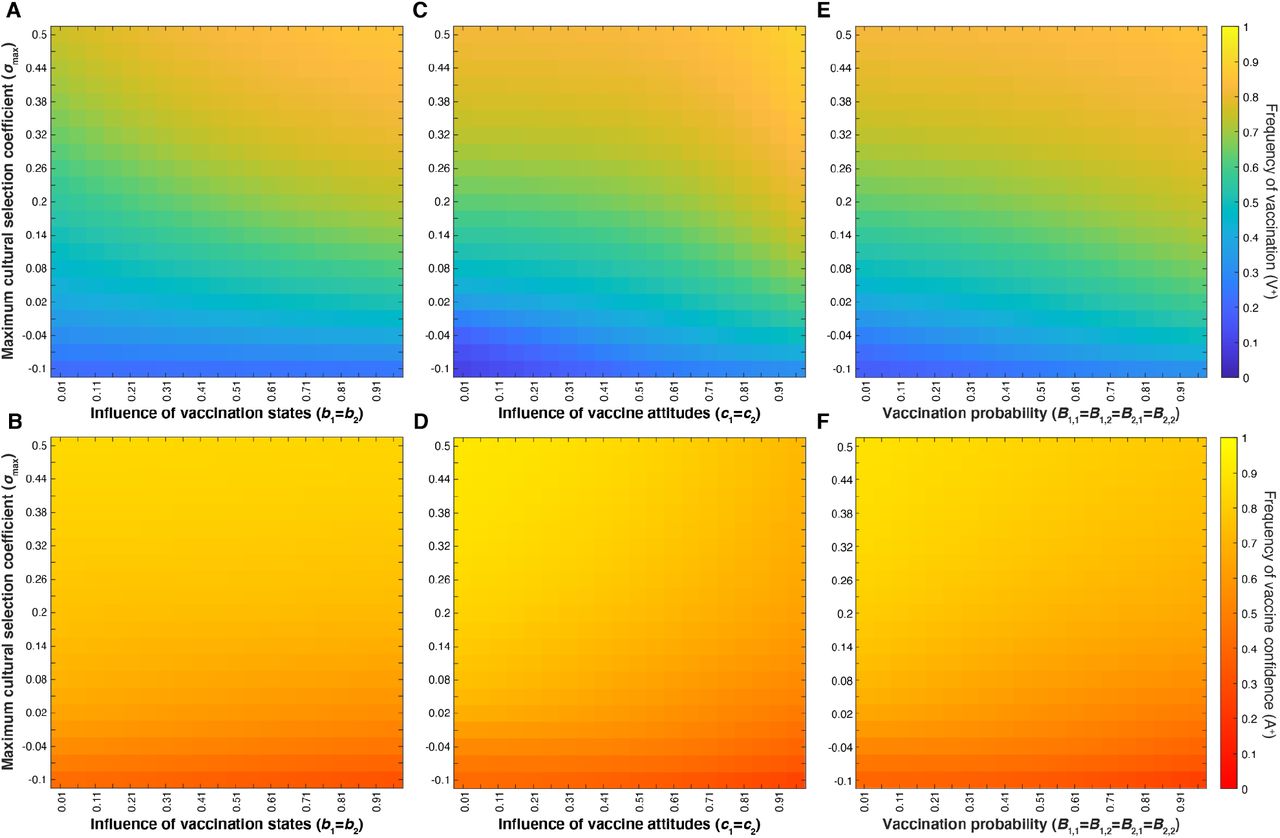
Heatmaps showing final vaccination coverage (A, C, E) and final vaccination confidence (B, D, F) after 100 time-steps with oblique transmission. As in previous figures, parameters not varied are given in Table 1. Parents’ likelihood of vaccinating their children depends on both their vaccination state and their attitude state. At default probabilities of vaccine confidence transmission (Cn), these figures show that modulating the maximum cultural selection coefficient affects the equilibrium levels of vaccination coverage and vaccine confidence across the range of specified parameters: parental vaccination state influence, bm (A, B), parental attitude state influence, cn (C,D), and offspring vaccination probability, Bm,n (E,F). Unless directly varied (as in panels E-F), Bm,n varies as bm and cn are varied, as shown in Table 1.
With neutral confidence transmission (C1 = C2 = 0.5), we also observe an expansion of the polymorphic space when we modulate cultural selection (σmax) alongside the influence and transmission parameters (Figure 11). Interestingly, in the cultural environment defined by this parameter space, we observe a pattern that deviates from the expected association between high vaccine confidence and high vaccination coverage: as the influence of vaccine attitudes (cn) and vaccination probabilities (Bm,n) increase (Figure 11, horizontal axes), the population’s equilibrium vaccination coverage increases while its vaccine confidence decreases. This pattern persisted across all tested levels of maximum cultural selection (σmax) (Figure 11, vertical axes). In other words, we observe higher levels of confidence at low influence and transmission probabilities than we do at higher probabilities (Figure 11D, F).
We explored the interaction between the influence parameters, bm and cn, at various maximum cultural selection coefficients (σmax) (Figure 12). Vaccination coverage and vaccine confidence equilibrate at mid-range frequencies (between 0.3 and 0.8) across the range of bm and cn, indicating that these trait frequencies are not particularly sensitive to either parameter.
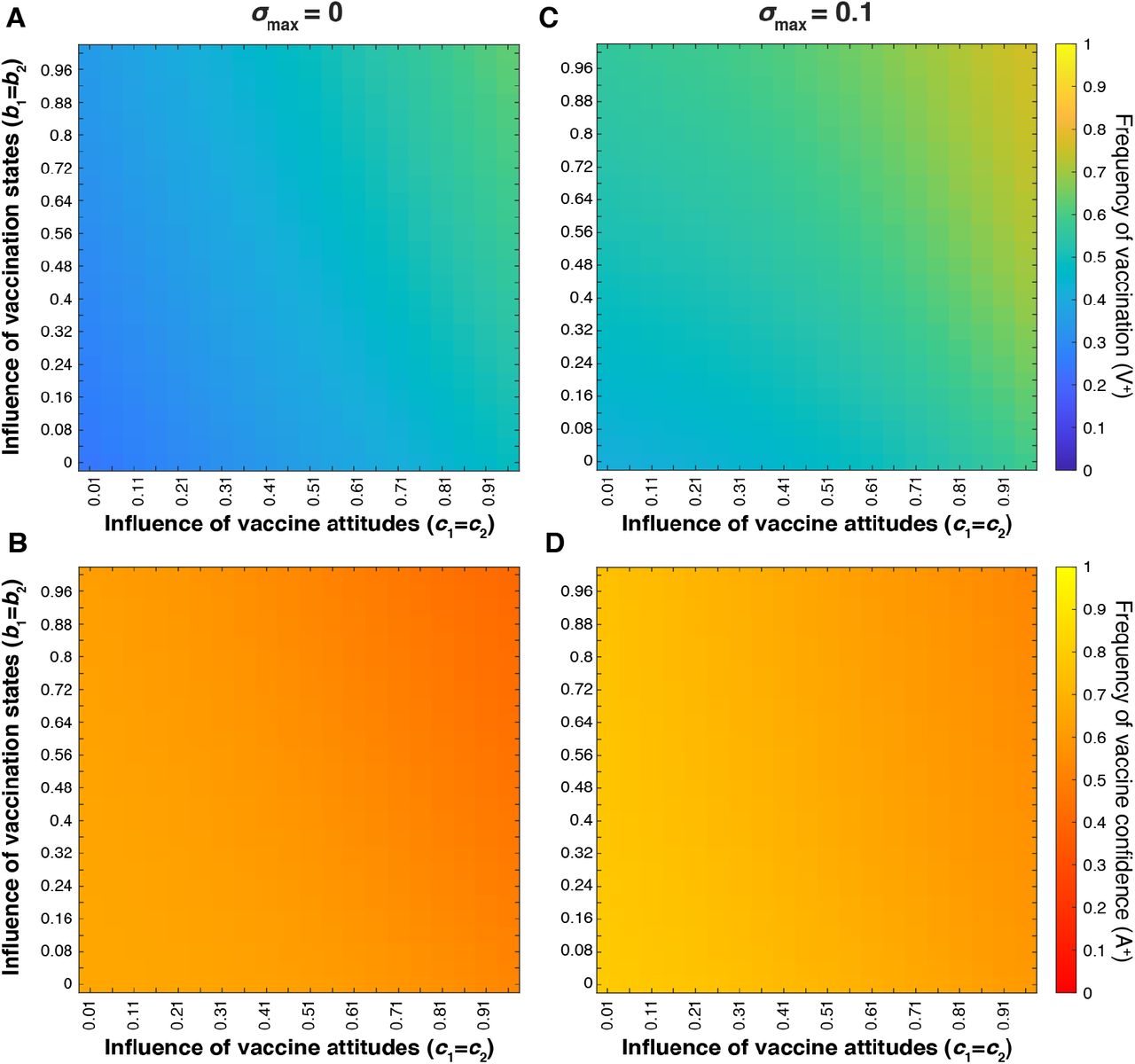
Heatmaps showing final vaccination coverage (A, C) and final vaccination confidence (B, D) after 100 timesteps with oblique transmission. We modulate the interaction between vaccination state influence (bm; vertical axis) and attitude influence (cn; horizontal axis) at various maximum cultural selection coefficients: σmax = 0 (A, B) and σmax = 0.1 (C, D). As in previous figures, unvaried parameters are given in Table 1. Vaccination frequency increases as both influence probabilities increase and vaccination confidence decreases as both influence probabilities increase.
Cultural selection favoring vaccination increases the equilibrium level of vaccination coverage and vaccine confidence (Figure 12 and Figure S7). The most notable deviation between equilibrium confidence and vaccination frequencies occurs at the intersection of the highest influence parameter values (bm and cn), circumstances in which the parents’ vaccination states and vaccine attitudes overwhelmingly support offspring vaccination; in this top right section of the heat maps, vaccination coverage is high while vaccine confidence is lower, indicating a behavioral pattern in which mixed-trait couples are more inclined to vaccinate their offspring than transmit vaccine confidence. Overall, the addition of oblique transmission to a population that would otherwise equilibrate at high vaccination coverage (Figure S2) leads to increased attitude transition to vaccine hesitancy and subsequently lower vaccine coverage.
Mating Preferences
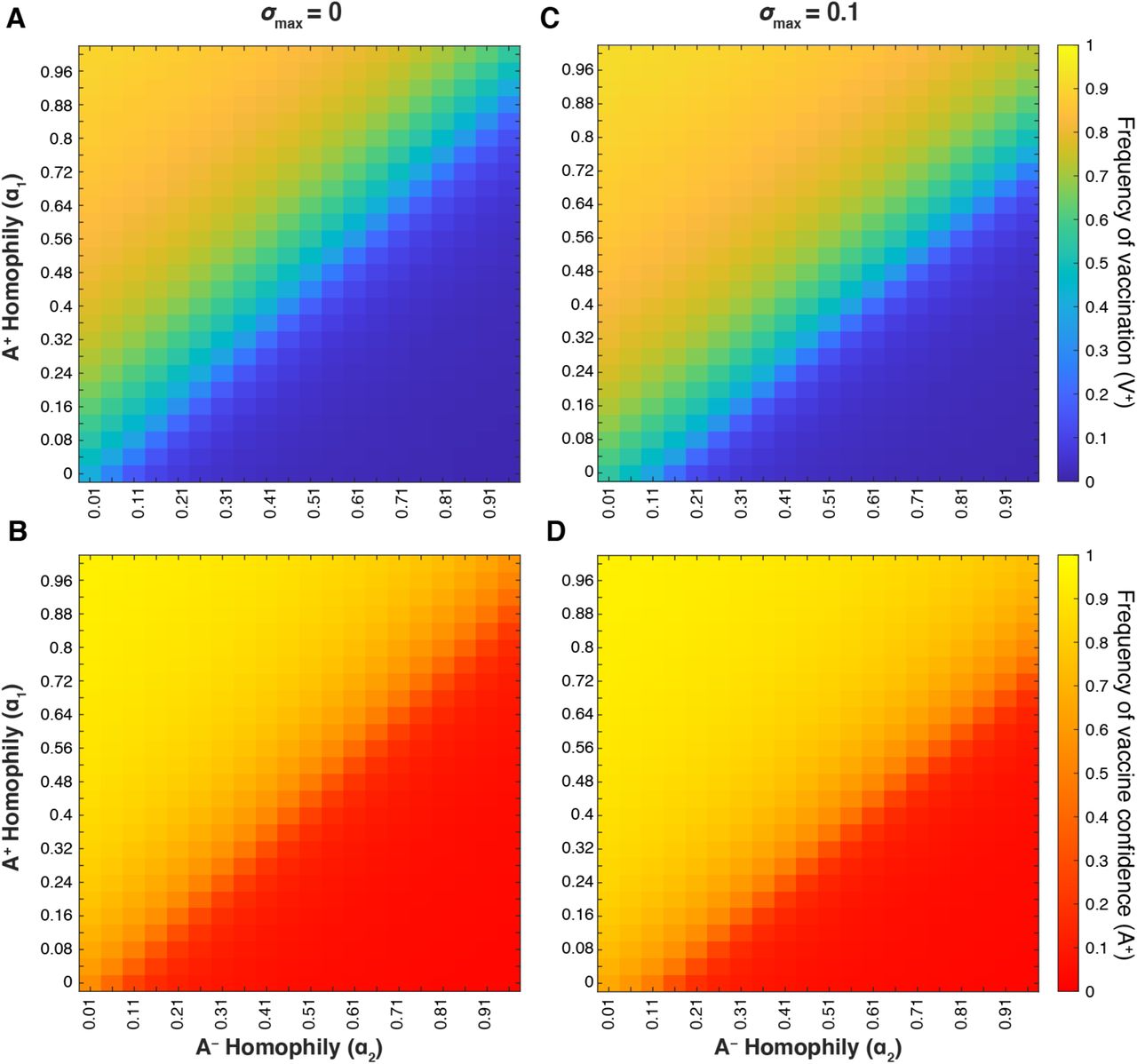
We hypothesized that mating preference (assortative mating) could modulate belief and behavior dynamics and thus the vaccination coverage and confidence levels in the population. If individuals are more likely to pair with individuals of the same vaccine attitude, such that same-attitude couples become more common and mixed-attitude couples are less common, the parameter values for mixed-attitude couples may have less impact on vaccination coverage and confidence dynamics. Therefore, we analyzed the interaction between A+ homophily (with α1 indicating the preference of A+ individuals for other A+ individuals) and A− homophily (with α2 indicating the preference of A− individuals for other A− individuals) at various σmax levels. When vaccine attitudes are transmitted both from parent to offspring and between unrelated individuals (vertical and oblique transmission) and there is neither cultural selection for nor against being vaccinated (σmax = 0), we observe a threshold region at roughly equal mating preferences (α1 ≈ α2; diagonal lines in Figure 13A-B); above this boundary (when α1 > α2) vaccination coverage and confidence are much higher than below this boundary (when α1 < α2). When cultural selection favors being vaccinated (σmax > 0, Figure 13C-D, Figure S8C-D), the threshold between high and low equilibrium values is shifted, such that high coverage and high confidence levels can potentially be attained even when vaccine hesitant individuals preferentially pair with each other more than vaccine confident individuals do (α1 < α2). Likewise, if cultural selection explicitly does not favor vaccination (e.g. σmax = −0.1, Figure S8), low vaccination coverage and confidence can occur even when there are more vaccine confident couples in the population than hesitant couples (α1 > α2). We observe qualitatively similar patterns when vaccine attitudes are only transmitted from parent to offspring (Figure S9); as we have previously observed, the addition of oblique transmission leads to a broader polymorphic region than vertical transmission alone. These patterns illustrate two overarching themes: 1) preferential interactions between individuals with similar vaccine beliefs can dramatically shift the equilibrium levels of vaccination coverage and confidence with all other parameters remaining equal, and 2) the actual and perceived quality and efficacy of the vaccine are important to determining vaccine acceptance.
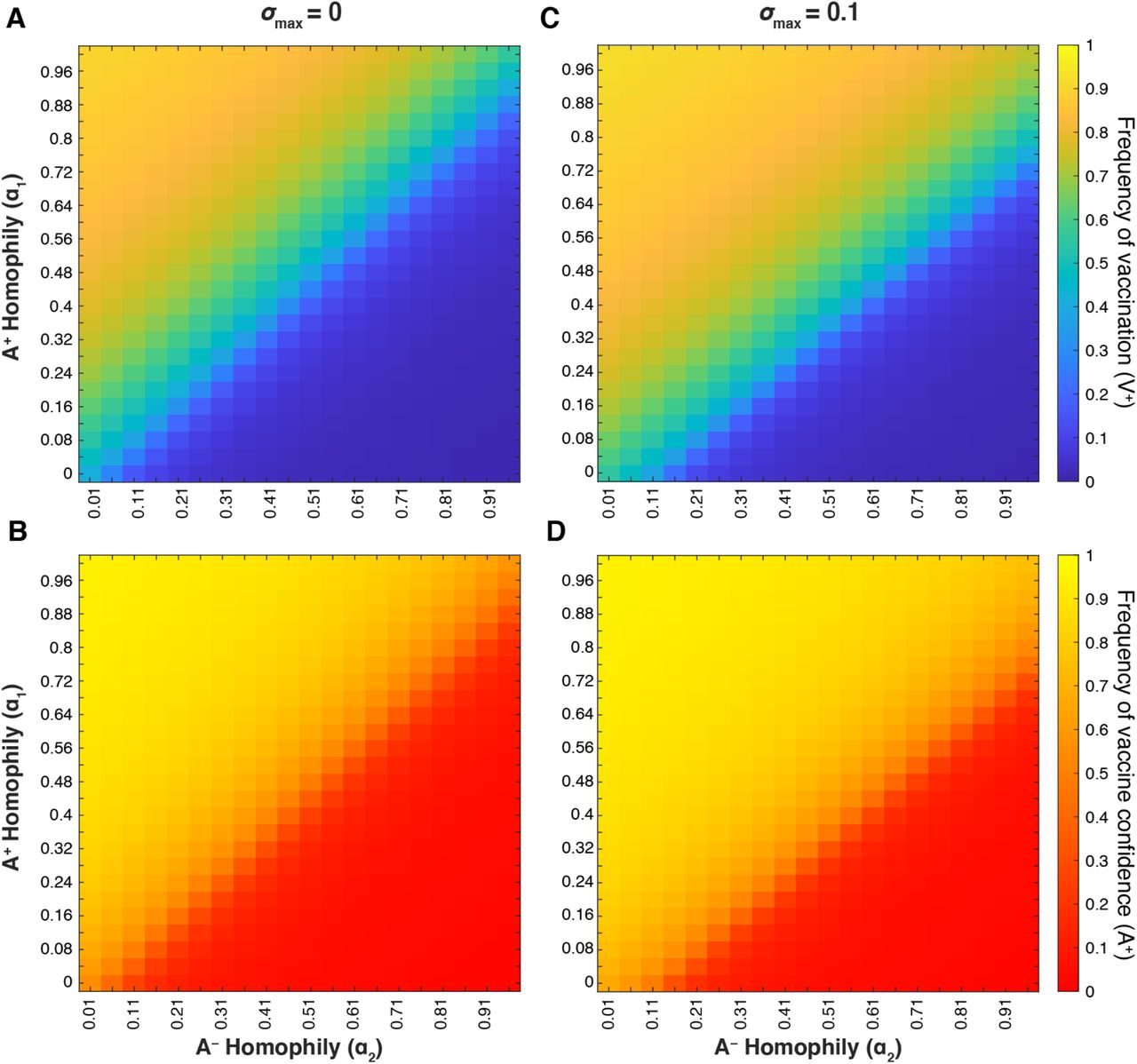
Heatmaps showing final vaccination coverage (A, C) and final vaccine confidence (B, D) after 100 timesteps with oblique transmission. As in previous figures, unspecified parameters are given in Table 1. As vaccine-hesitant individuals (A−) increasingly prefer to pair with one another (α2; horizontal axis), vaccine-confident individuals (A+) must also preferentially interact to maintain high vaccine coverage (α1; vertical axis); this tradeoff is modulated by the cultural selection pressures on vaccination (σmax = 0 (A, B) and σmax = 0.1 (C, D)).
Mandatory Vaccines and Vaccine Inaccessibility
Compulsory Vaccination
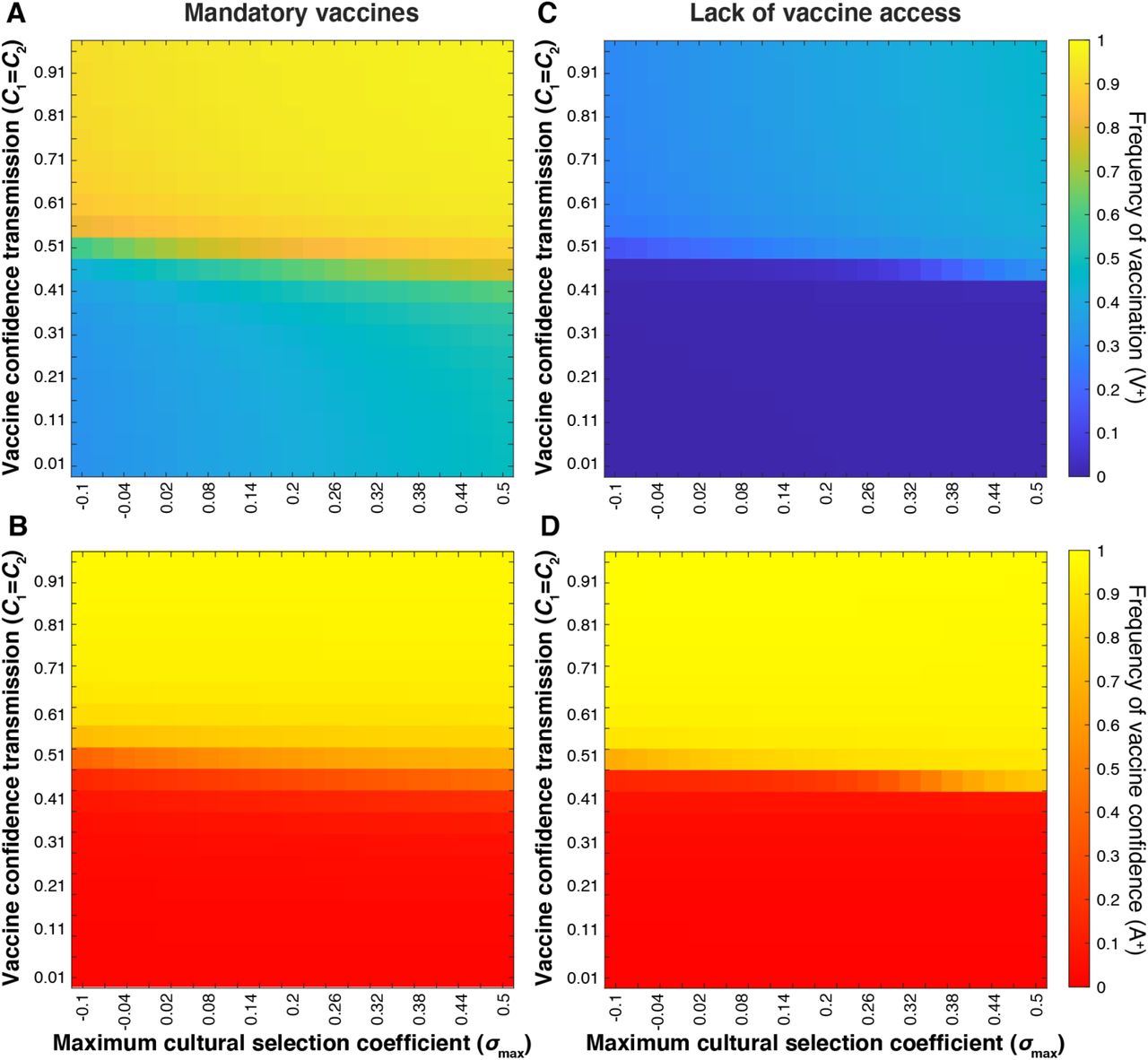
The implementation of mandatory childhood vaccinations has resulted in almost 100% vaccination coverage in the United States [9,61]. Allowing non-medical exemptions such as those based on philosophical or religious beliefs, however, has contributed to the recent decrease in vaccination rates [64]. We suspect that parental vaccine attitudes also influence their use of exemptions and thus levels of non-vaccination under a mandated vaccination system. Therefore, we simulate the effects of varying levels of compulsory vaccine mandate strictness (i.e. the ease of obtaining an exemption) by modulating the influence of vaccine attitude on offspring vaccination for each pairing. We assume the implementation of strict mandates (difficulty in obtaining exemptions) would increase vaccination in all couple types. If vaccination exemptions are permitted, we expect that A− × A− couples (those with two vaccine-hesitant individuals) would be most likely to obtain exemptions, followed by mixed attitude (A− × A+ or A+ × A−) couples, with vaccine confident couples (A+ × A+) being least likely. Hence, to model the effects of implementing a strict mandate, we increase attitude influence parameters from baseline values (Table 1) to c0 = 0.5, c1 = c2 = 0.9, c3 = 0.99, then examined the effect of cultural selection coefficient and confidence transmission probability (Figure 14A-B); we then compare the results of this strict mandate both to our baseline parameters and to a more lenient mandate represented by c0 = 0.3, c1 = c2 = 0.7, c3 = 0.99 (Figure S10).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
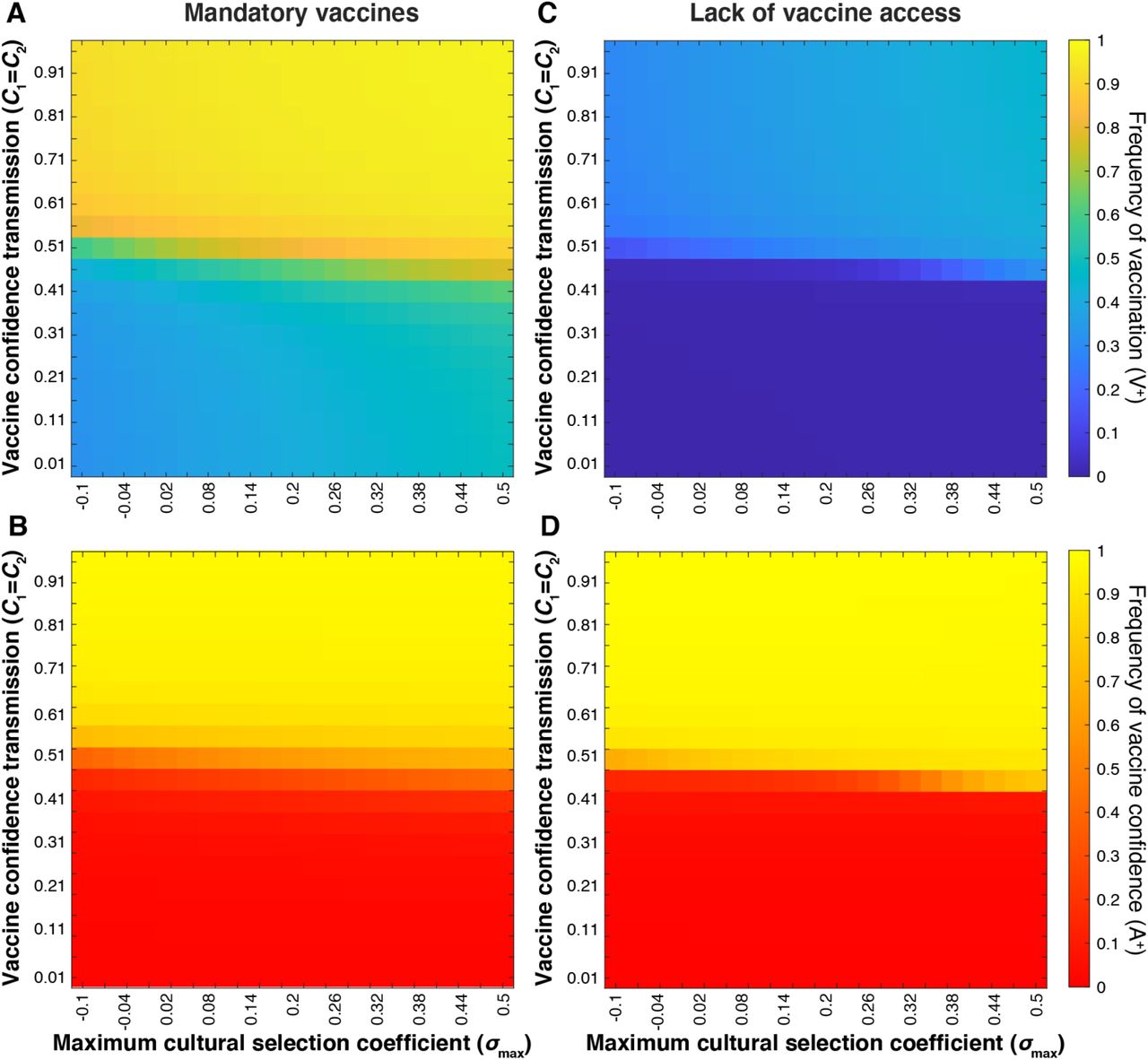
Equilibrium vaccine coverage and vaccine confidence levels for varying values of confidence transmission (Cn; vertical axis) and maximum selection coefficient (σmax; horizontal axis). A strict vaccine mandate (A, B) is simulated by c0 = 0.5, c1 = c2 = 0.9, c3 = 0.99; Vaccine scarcity (C, D) is simulated by c0 = 0.01; c1 = c2 = 0.1, and c3 = 0.5. As in previous figures, unspecified parameters are given in Table 1. These simulations show an inverse correlation between vaccination coverage and vaccine confidence at Cn < 0.5 under a strict mandate, and Cn > 0.5 when vaccine access is severely limited.
Our previous simulations consistently showed a positive correlation between population-level frequencies of vaccination coverage and vaccine confidence (Figures 8–10, 13). However, modeling a cultural environment under strict mandates reveals a decoupling of vaccination coverage and vaccine confidence at low confidence transmission probabilities (Figure 14A-B).
Even when vaccine confidence is very low (specifically at mixed-trait couple confidence transmission probabilities below 0.5; red region in Figure 14B), vaccination coverage is much higher than without the mandate (compare Figure 14A-B to Figure 10A-B, see also Figure S10). This suggests that an external pressure to vaccinate is able to overcome the opposing cultural pressure imposed by hesitancy in the population. If we lower the barrier to acquiring an exemption (using more lenient mandate parameters), vaccination coverage and vaccine confidence dynamics begin to mirror one another, as they do in simulations without vaccine mandates (Figure S10).
Vaccine Inaccessibility
We next modeled the dynamics of vaccination behaviors and attitudes when a population has limited access to vaccines. To represent this scenario, we reduced the influence of parental vaccine attitudes on vaccination behaviors for couples with at least one confident individual (i.e. reducing c1, c2, c3 from default values). In this simple representation of a vaccine-scarce environment, we assume that parents’ confidence in vaccines would have reduced influence on their ability to vaccinate their offspring, that is, their vaccine confidence does not ensure their ability to overcome vaccine inaccessibility. Hesitant couples are least likely to vaccinate their offspring regardless of vaccine availability, but couples who would likely vaccinate their offspring given the chance would have difficulty doing so due to the lack of resources. This scenario can be contrasted with the strict vaccination mandate scenario described above, in which a couple would often vaccinate their children despite their hesitancy. Attitude influence parameters were set to c0 = 0.01; c1 = c2 = 0.1, and c3 = 0.5, and as before, we modulated the maximum cultural selection coefficient of vaccination (σmax) and confidence transmission (C1 and C2) (Figure 14C-D). Vaccination coverage was noticeably reduced overall, while vaccine confidence increased slightly across the parameter space. Juxtaposed with the strict mandate (Figure 14A-B), our vaccine scarcity models produce an opposite deviation of vaccination coverage from vaccine confidence levels: when vaccines are mandated, we observe increased vaccination coverage in low-confidence environments, and when vaccines are inaccessible, we observe lower than expected vaccination coverage (<50%) in a predominantly vaccine-confident environment (>90%) (Figure 14).
Discussion
In this manuscript, we present a simplified model of a complex process: the spread of vaccine attitudes and their effects on vaccination behavior in a population. Increasing and maintaining sufficient vaccination coverage to combat disease is more complex than simply increasing vaccine availability or providing accurate information. A number of factors affect a person’s vaccine-related beliefs and a family’s decision to vaccinate their children, including their history with vaccinations and their perception of the disease and vaccine effects. As such, it is important that we understand how these personal factors can shape vaccination cultures and thus affect public health. Using a cultural niche construction framework, we modeled the spread of vaccine attitudes and vaccination behavior in a variety of circumstances and measured the resulting levels of vaccination coverage and vaccine confidence in the modeled population. Using this novel approach of modeling dynamically interacting beliefs and behaviors, we are able to explore the interplay of cultural factors that drive vaccine attitudes and vaccination behavior, providing insight into how vaccination cultures are formed, maintained, and evolve.
Our model shows vaccine confidence transmission (Cn) to be the parameter that most strongly determines vaccination coverage and confidence levels. That is, the probability of transmitting vaccine-positive attitudes is a strong predictor of future vaccination coverage (Figures 4 and 10). We note, however, that the vaccine confidence transmission of mixed-trait couples (those with one hesitant parent and one confident parent, C1 = C2) has a much larger effect on the levels of vaccination coverage than mixed-trait couple vaccination probability itself, likely because the spread of vaccine confidence or hesitancy makes mixed-trait couples relatively rare in the population. By modulating the vaccination probabilities of all individuals directly (varying all Bm,n), while modulating mixed-trait confidence transmission (C1 = C2) (Figure 5) or all confidence transmission parameters (Cn) (Figure S1), we show that vaccination probabilities work alongside vaccine confidence transmission to determine vaccination coverage equilibria, while confidence equilibria are dictated primarily by confidence transmission probabilities.
It is important to note that even though this model demonstrates the powerful effects of beliefs on vaccination behaviors, it also shows that vaccine efficacy and perceived value are important to maintaining sufficient levels of vaccination coverage, especially if vaccine confidence is not being robustly transmitted (or maintained in adulthood). Further, we show that a culture in which vaccine-hesitant individuals preferentially assort with one another more so than vaccine-confident individuals (α2 > α1) can allow vaccine hesitancy to more easily gain a foothold, lowering vaccination coverage unless vaccines have a high perceived value (i.e. high σmax, Figure 13, Figure S9). Thus, our simulations suggest that a pro-vaccination health culture can be undermined by a vaccine-hesitancy “echo chamber”. This phenomenon would likely be amplified if we modeled homophily of oblique interactions as well, for example if individuals preferentially interacted with friend groups or news sources that shared similar beliefs and values to their own.
Further, our model demonstrates that the equilibrium frequencies of vaccination coverage and vaccine confidence are determined by the cultural context rather than by the initial frequencies. In other words, the current cultural landscape—beliefs, behaviors, policies, etc.—are more predictive of future levels of vaccine coverage and confidence than current coverage and confidence levels in the population. This result further supports the importance of considering the cultural factors that have shaped current health-related beliefs and behaviors if health policies aim to maintain or change the current conditions.
In the absence of external pressures, populations reach an equilibrium at which vaccination coverage and vaccine confidence mirror each other across the tested parameters: high vaccination coverage coincides with high levels of confidence in vaccines, and low coverage coincides with low confidence levels. This result suggests, intuitively, that when population traits are at or near an equilibrium, we can infer that a population with high vaccination coverage will have low rates of vaccine hesitancy and vice versa. We then model two possible exceptions to this pattern—vaccine mandates and a lack of access to vaccines. When there is increased pressure to vaccinate or difficulty in acquiring vaccination exemptions, an undercurrent of vaccine hesitancy can persist in a relatively well-vaccinated population, potentially limiting the adoption of newly introduced vaccines. This possibly contributes to the unexpected lag in uptake of newer vaccines, such as the COVID or HPV vaccines, in communities with otherwise high vaccination rates [65–67]. The perceived increase in hesitancy surrounding new vaccines may actually be existing vaccine hesitancy becoming evident. In addition, “fence sitters”, those who have not made a firm stance regarding vaccines and thus could be more influenced by targeted campaigns [61], may develop higher levels of uncertainty about new vaccines than their parents had about existing ones.
In contrast to the effect of vaccine mandates, by modeling vaccine inaccessibility we illustrate another important pattern: reduced vaccination coverage in a vaccine confident culture. In a vaccine-scarce environment, an individual’s attitude regarding vaccines has less influence on vaccination behavior due to the barrier imposed by resource availability. As a result, a population may be undervaccinated despite holding vaccine-affirming beliefs. In addition, a health culture previously shaped by vaccine inaccessibility could potentially ingrain specific behavioral practices (for example, visiting the doctor only when a child is sick and not for a regular vaccine schedule) that are not easily modified even if vaccines become more readily available. These vaccine scarcity scenarios are most likely to exist in low-and middle-income countries in which vaccine acquisition, storage and/or distribution resources are insufficient [68–70] whereas the opposite scenario (low confidence/high vaccination) after vaccine mandates is most common in developed nations [71]. In summary, we find that vaccine mandates can result in high vaccination coverage even in a culture of hesitancy, and that lack of access to vaccines can produce the inverse: low vaccination coverage in a culture of confidence.
Individuals consider multiple factors alongside vaccine efficacy when deciding whether to vaccinate themselves or their children, essentially performing an internal cost-benefit analysis based on their circumstances and interpretation of accessible information. We aimed to be inclusive of these considerations via our comprehensive cultural selection coefficient; this parameter allows the frequency of vaccination to deviate from the level that is expected given the cultural transmission probabilities. Increasing positive public perception through honest and effective communication, while reducing public concern about vaccines and increasing vaccine safety, together could drive increased vaccination trust and acceptance. Achieving the optimal vaccination coverage lies not only in the hands of the public by vaccinating themselves and their children, but also in the efforts of health officials and leaders in creating an environment that fosters confidence by assuring the public of vaccine efficacy, safety, and value, while providing convenient avenues to attain vaccines.
As with any model, we cannot fully capture the complex reality of vaccine hesitancy and vaccination behavior. First, though vaccination frequency data is available for numerous vaccines, frequency data for vaccine attitudes are much less common, with the two traditionally not surveyed together. Thus, there is no dataset that exactly estimates the phenotypes presented here, for example, the number of vaccinated but hesitant (V+A−) individuals in a population. The goals of vaccination attitude surveys have been primarily to identify themes of vaccine hesitancy, and to a lesser degree, the themes of vaccination. However, they do not report parent vaccination states or whether the child was actually vaccinated (on schedule). With data presenting parent vaccination states alongside their vaccine attitudes and vaccination decisions, we would be able to more accurately inform phenotype frequencies, possibly extending the model to incorporate various types of hesitancy. We note, however, that our results did not depend on the initial proportions of vaccination status or vaccine hesitancy, so these data would primarily be for comparison to our equilibrium outcomes. We were also constrained by limited data to inform the cultural transmission and transition probabilities. In our model, baseline confidence transmission and influence probabilities are structured according to a simple pattern of inheritance, such that each parent is equally likely to influence an offspring’s phenotype.
However, cultural traits and vaccination attitudes may not strictly follow this pattern: one parent might have more influence, or one variant of a trait might be more likely to be transmitted. In addition, transmission probabilities are constant in our model, remaining unaffected by changing cultural conditions throughout each simulation, but in reality, these probabilities may fluctuate in response to a variety of factors including vaccine type or family structure. Future developments of the model could include modulating the probability of transmission of vaccine confidence dependent on aspects of the cultural environment, such as the attitude frequencies in the population. Our cultural selection coefficient and attitude transition probabilities did vary with the frequency of vaccination coverage, but the exact relationships could not be informed by existing data. As a result, we generated frequency-dependent functions to fit a set of assumptions: for example, that the transition to vaccine hesitancy might be most common when the vaccination coverage is low and the benefits of vaccination are more apparent.
Though vaccination coverage and vaccine confidence stabilized (reached equilibria) in our simulations, in reality vaccination rates fluctuate over time in response to changing population dynamics, perhaps never arriving at a stable equilibrium. For example, the increasingly rapid spread of information [23] may cause attitudes and behaviors to change frequently over short periods of time. In our model, most of the phenotype frequency fluctuations occur in the first few iterations before quickly adjusting to an equilibrium. Unlike many models of population dynamics, this model has a discrete-time format and does not consider a birth-death cycle or asynchrony in population turnover. Thus, the timescale of our model might not translate directly to years or generations, and we avoid interpreting the number of iterations in literal terms. It is possible that if more realistic birth and death processes were incorporated, the cultural dynamics would occur at different timescales and would continue to fluctuate instead of approaching a stable equilibrium. In addition, parents’ vaccination decisions are also influenced by the grandparents of the children to be vaccinated [72]. A restructuring of the timescale or the incorporation of population asynchrony in our model could allow for consideration of these influences.
Finally, we constructed the offspring vaccination probability to be informed primarily by parents’ vaccine attitudes and secondarily by their own vaccination status. Though it is understood that there is an interaction between parents’ beliefs and their own experiences with vaccines regarding their decision to vaccinate their children, accurately modeling the relative contribution of these two factors could benefit from empirical studies on parental willingness to vaccinate based on their beliefs and vaccination status. With our current formula (Bm,n, Table S2), vaccine-confident parents who did not themselves receive childhood vaccines have a reduced likelihood of vaccinating their offspring than vaccinated parents. In reality, parental vaccine attitudes might even further outweigh their own vaccination status in their decision making than we model here.
Our findings, which are based on cultural evolutionary modeling in a public-health context, suggest several avenues for policy and outreach recommendations. First, we note that vaccine mandates with limited exemptions will increase the vaccination coverage but may mask the spread of vaccine hesitancy. In addition to vaccine policy, our results suggest that a broad effort to encourage and inform the public about vaccine safety and efficacy will foster higher vaccine coverage. In this vein, we note that individuals who are skeptical about vaccines might invest more time in seeking out information about them [73–75], thus we recommend that accurate information about vaccines should be readily accessible through a variety of means, be easily understood, and be supported by personal anecdotes. If misinformation is easier to encounter and digest than accurate information, it could have an outsized impact on individuals who are “on the fence” [61]. Relatedly, we note that research suggests transparency and trust go hand in hand: if individuals perceive the healthcare system is concealing information to make vaccines appear more safe, trust in that system will decrease and people will be more susceptible to vaccine hesitancy [73–76]. Finally, dialogue between people with different beliefs and attitudes can help to break the “echo chambers” of homophily, encouraging individuals to communicate and empathize with one another. Therefore, to address vaccine hesitancy, our results underscore the importance of considering the cultural beliefs that underpin health behaviors.
References




Subject Area
Reviews and Context
0
Comment
0
TRIP Peer Reviews
0
Community Reviews
0
Automated Services
0
Blogs/Media
Author Videos