Article Text

Abstract
Objective The early intestinal microbiota exerts important stimuli for immune development, and a reduced microbial exposure as well as caesarean section (CS) has been associated with the development of allergic disease. Here we address how microbiota development in infants is affected by mode of delivery, and relate differences in colonisation patterns to the maturation of a balanced Th1/Th2 immune response.
Design The postnatal intestinal colonisation pattern was investigated in 24 infants, born vaginally (15) or by CS (nine). The intestinal microbiota were characterised using pyrosequencing of 16S rRNA genes at 1 week and 1, 3, 6, 12 and 24 months after birth. Venous blood levels of Th1- and Th2-associated chemokines were measured at 6, 12 and 24 months.
Results Infants born through CS had lower total microbiota diversity during the first 2 years of life. CS delivered infants also had a lower abundance and diversity of the Bacteroidetes phylum and were less often colonised with the Bacteroidetes phylum. Infants born through CS had significantly lower levels of the Th1-associated chemokines CXCL10 and CXCL11 in blood.
Conclusions CS was associated with a lower total microbial diversity, delayed colonisation of the Bacteroidetes phylum and reduced Th1 responses during the first 2 years of life.
- Chemokines
- Infant Gut
- Intestinal Bacteria
- Intestinal Microbiology
- Molecular Biology
Statistics from Altmetric.com
Significance of this study
What is already known on this subject?
-
The infant gut microbiota diversity increases during the first years of life.
-
The microbiota composition differs between infants born by caesarean section (CS) or vaginal delivery with a delayed colonisation of the genus Bacteroides.
-
Bacterial colonisation is necessary for the development of the immune system and immune regulation.
-
An association between CS delivery and the development of allergic disease has been observed in several studies.
What are the new findings?
-
The total microbiota diversity is lower in CS than vaginally delivered infants through the first 2 years of life.
-
The diversity of the Bacteroidetes phylum is lower in CS born infants during the first 2 years of life.
-
Vaginal delivery is associated with increased circulating levels of Th1-associated chemokines during infancy.
How might it impact on clinical practice in the foreseeable future?
-
Deeper knowledge of the impact of delivery mode on microbiota composition and immune regulation may lead to improved allergy preventive strategies.
Introduction
The gastrointestinal tract of the newborn infant is considered to be sterile. Bacteria from the environment, mainly from the mother, colonise the infant gut immediately following birth. Dominant members of anaerobic Firmicutes and Bacteroidetes do not appear to grow outside the gut and hence need to be transmitted between human hosts.1 To what extent the transmission occurs from mother to offspring is not clear, but differences in microbiota composition depending on delivery mode indicate a mother–child transmission during vaginal delivery. A recent study based on pyrosequencing of 16S rRNA genes demonstrated that the microbiota of vaginally delivered (VD) neonates (<24 h postdelivery) resembled the vaginal microbiota of their own mother and was similar across multiple body habitats (skin, oral, nasopharynx and faeces), while in neonates born by caesarean section (CS), it resembled the mother's skin microbiota.2 While this study provided evidence that microbiota from the birth canal is transferred from mother to child, providing an inoculum for the initial microbiota, it remains to be shown that specific gut microbes are successfully transmitted during vaginal delivery.
The incidence of caesarean delivery has increased from 5% in the 1970 s to more than 60% in some hospitals in China according to recent reports.3 The early colonisation pattern differs between VD infants and those delivered by CS, including a delayed colonisation of, for example, Bacteroides and Bifidobacterium spp. in CS infants.4 ,5 The influence of delivery mode on gut microbiota development has not previously been longitudinally characterised using powerful cultivation-independent microbiological methods, however. The intestinal microbiota is important for the development of the immune system and immunological tolerance.6 Differences in the postnatal microbial colonisation may explain the higher incidence of immune mediated diseases such as allergy in children born by CS as compared with those born vaginally.7 ,8 Indeed, allergic disease has been associated with a low prevalence of Bacteroides and Bifidobacterium,9 ,10 and a low intestinal microbiota biodiversity in early infancy appears to have an impact on the development of allergic disease later in life.11–13 A failure of Th2-silencing during maturation of the immune system may underlie development of Th2-mediated allergic disease.14 Appropriate microbial stimulation may be required to avoid this pathophysiological process, as early differences in the gut microbiota may shape later immune development.6 ,15 The influence of CS on immune development is largely unknown, however.16 The aim of the present study was to monitor the development of the infant intestinal microbiota in babies born vaginally and through CS, and to relate the findings to the maternal microbiota and to Th1- and Th2-associated chemokine levels during infancy.
Materials and methods
Ethics
The human ethic committee at Linköping University, Linköping, Sweden, approved the study. Informed consent was obtained from both parents before inclusion.
Subjects and sample collection
The study group comprised 24 healthy women and their infants. Nine of the infants were born by CS and 15 by VD (see online supplementary table S1 regarding, sex, delivery, birth weight, the use of antimicrobials and length of breast feeding for the different infants). Seven out of nine mothers, who gave birth through CS, were given antibiotics prophylactically during the surgery (see online supplementary table S2). This was done after the delivery, however, and thus the infants were not exposed to antibiotics via the placenta. No children were treated with antibiotics during the neonatal period. Twenty of the infants (83%) were partly breast fed until at least 6 months of age. The women and children included in this study were part of a larger study assessing the prevention of allergic disease by probiotics17 (ClinicalTrials.gov ID NCT01285830) and they all received placebo. Stool samples were collected from the mothers 1 week after delivery and from the children at 1 week, 1, 3, 6, 12 and 24 months. The faecal samples were immediately frozen at −20°C following collections and later stored at −70°C. Samples were collected in 2002–2005 and stored in −70°C until DNA extractions were conducted in 2009. No systematic differences in storage times existed between the VD and CS samples (mean months: VD 80±7, CS 78±4, Student t test, p=0.42).
DNA extraction and 16S rRNA gene amplification
The DNA extraction and 16S rRNA gene amplification were performed as described previously18 with the following modifications; the primer pair used, targeting the variable regions 3 and 4 of the 16S rRNA gene, was 341f 5′CCTACGGGNGGCWGCAG with adaptor B and 805r 5′GACTACHVGGGTATCTAATCC with adaptor A and a sample-specific barcode sequence consisting of five nucleotides.19 The barcodes contained no homopolymers and a pair of barcodes differed in at least two positions. The 341f-805r primer pair was shown to be the least biased among 512 primer pairs evaluated in silico for bacterial amplification and was experimentally shown to give a taxonomic composition similar to shotgun metagenomics.20 The primer pair has good coverage of bacterial groups typically found in the human lower intestine. For phylum Bacteroidetes, 121 862 of 132 120, for phylum Firmicutes 376 912 of 406 649, for phylum Proteobacteria 336 471 of 368 375 and for family Bifidobacteriaceae 1112 of the 1239 sequences are matched, when considering Ribosomal Database Project (RDP) sequences that span the region. A negative PCR reaction without template was also included for all primer pairs in each run. The PCR products with approximate lengths of 450 bp were purified with AMPure beads (Becton Dickinson, Franklin Lakes, New Jersey, USA) using a Magnet Particle Separator (Invitrogen, Carlsbad, California, USA). The concentrations of the purified products were measured by Qubit fluorometer (Invitrogen), the quality was assessed on a Bioanalyzer 2100 (Agilent, Santa Clara, California, USA), and the samples were amplified in PCR-mixture-in-oil emulsions and sequenced from the 805r primer on different lanes of a 2-lane PicoTiterPlate on a Genome Sequencer FLX system (Roche, Basel, Switzerland) at the Swedish Institute for Communicable Disease Control (Solna, Sweden). Sequence processing was carried out with the AmpliconNoise software package21 correcting for errors introduced in the PCR and pyrosequencing as well as removing chimeric sequences. Also, reads lacking a correct primer and/or having less than 360 successful pyrosequencing flows were excluded.21 Denoised sequences were trimmed to 198 bp after primer and barcode removal and clustered by complete linkage clustering into operational taxonomic units (OTUs) at the 97% similarity level using AmpliconNoise.
Taxonomic classification
Each denoised sequence, as well as the most abundant sequence for each OTU, was BLAST searched with default parameters against a local BLAST database comprising 836 814 near full-length bacterial 16S rRNA gene sequences from the RDP v.10.10.22 The sequences inherited the taxonomic annotation (down to genus level) of the best scoring RDP hit fulfilling the criteria of ≥95% identity over an alignment of length ≥180 bp. If no such hit was found the sequence was classified as ‘no match’. If multiple best hits (of same score) were found, the taxonomy was set to the most detailed level of taxonomy shared by the best hits. The majority of reads had an RDP relative within 95% sequence similarity and were hence of bacterial origin. After removal of pyrosequencing noise and chimeric sequences using the AmpliconNoise package,21 357 685 high quality, typically 198 bp long, sequence reads remained, with 828–4395 reads per sample (mean=2129). These corresponded to 3048 unique sequences and 1818 OTUs, clustered at 97% similarity level using complete linkage clustering.
Sample clustering
The online version of Fast Unifrac (http://bmf2.colorado.edu/fastunifrac/)23 was used to calculate weighted sample distances by mapping our OTU sequences with BLAST onto the Greengenes reference sequences (downloaded from the Fast Unifrac web page, May 2009) and using the corresponding Greengenes tree. A principal coordinates analysis plot based on all pair-wise sample distance was created on the Fast Unifrac web page. Our OTU sequences were mapped onto 154 Greengenes sequences.
Statistical testing for over-representation or under-representation of bacterial lineages
Statistical tests of over-representation or under-representation of bacterial lineages among sample groups were made at the phylum and genus levels using Wilcoxon rank-sum test. To correct for multiple testing, the p values were converted to false discovery rate values (Q values).
Diversity estimations
Shannon diversity index was calculated as H=–Σ log(pi)pi, where pi denotes the relative frequency of OTU i,24 Pielou's evenness index as –Σ log(pi)pi/log(Sobs), where Sobs denotes the number of observed OTUs in the sample,25 and Chao1 richness estimate as Sobs + n1(n1 − 1)/n2(n2 − 1), where n1 and n2 are the number of observed singleton and doubleton OTUs, respectively.26 Since these parameters are influenced by sequencing depth and the sequencing depth differed between samples, we subsampled (with replacement) 1400 reads from each sample, counted the occurrences of the corresponding OTUs and performed the diversity calculations on these counts. This was repeated 10 times and averages of the diversity parameters calculated and used for further analysis. Four (out of 168) samples had fewer than 1400 reads and were excluded from this part of the analysis (three VD infants: one at 1 week, one at 3 months and one at 12 months, and one CS infant at 1 month). Diversity calculations and statistics were done with the R software (http://www.r-project.org/) and the R package vegan (http://cran.r-project.org/web/packages/vegan/). Repeated measures ANOVA were employed, using the Shannon diversity index at the different time points (1 week, 1, 3, 6, 12 and 24 months) as the repeated measures.
Statistical testing of mother–child overlap in sequence types
For each time point and for each infant, we calculated the number of specific sequences (sequence types) shared with its mother/number of sequence types observed in the infant (Rown). Likewise, we calculated the average number of sequence types shared with other mothers/number of sequence types observed in the infant (Rother). We then compared the Rown and Rother values pair-wise for all infants within each group (VD or CS) with the Wilcoxon signed rank test.
Chemokine analyses in venous blood and association with mode of delivery
Venous blood was collected at 6 (n=24), 12 (n=24) and 24 months (n=24) and stored as plasma or serum in −20°C pending analysis. The Th1-associated chemokines CXCL10 and CXCL11 and the Th2-associated chemokines CCL17 and CCL22 were analysed with an inhouse multiplexed Luminex assay.27 ,28 The limit of detection was 6 pg/mL for CXCL10 and CXCL11 and 2 pg/mL for CCL17 and for CCL22. All samples were analysed in duplicates and the sample was reanalysed if the coefficient of variance was >15%. The chemokine levels were compared between infants being born vaginally or by CS by repeated measures ANOVA using log transformed chemokine levels at the different time points (6, 12 and 24 months) as the repeated measures.
Results
Microbiota succession in infants
At the phylum level, the microbiota developed in a similar fashion in infants delivered vaginally and by CS, with a gradual decline in Proteobacteria from 1 week to 24 months, a peak of Actinobacteria at 3 months, an expansion of Firmicutes from 3 months and onward, and the emergence of Verrucomicrobia at around 6 months of age (figure 1, see online supplementary table S3). However, a notable difference between the VD and CS infants was the higher proportion of Bacteroidetes in VD infants during the first 12 months (significantly higher at 1 week, 3 months and 12 months; see online supplementary table S3).

Phylum level microbiota composition in mothers and their infants at 1 week, 1, 3, 6, 12 and 24 months. The mean relative abundances (%) of the most abundant bacterial phyla in the 15 vaginally delivered infants (A) and nine caesarean section infants (B) as well as in their mothers are shown.
The maternal microbiota resembled the typical adult flora as demonstrated in several previous studies18 ,29 ,30 and was independent of delivery mode. The Firmicutes was the dominating phylum, representing 74% and 71% in mean relative abundance for the 15 VD and nine CS mothers, respectively, followed by Bacteroidetes (16% and 13%, respectively), Actinobacteria (7% and 12%, respectively), Proteobacteria (3% and 2%, respectively) and Verrucomicrobia (1% and 2%, respectively) (figure 1, see online supplementary table S3).
The relative abundance of the major genera found in the infants and mothers are illustrated in figure 2A,B (see also online supplementary table S4). Infants in the VD group were colonised by Bacteroides to a greater extent than in the CS group (significantly more at 1 week (11/15 in VD vs 1/9 in CS; Fischer's exact test p=0.005), 3 months (11/15 vs 1/9; p=0.005) and 12 months (14/15 vs 4/9; p=0.015)). At 1 month of age, Bifidobacterium dominated the microbiota in both groups. The Enterococcus genus was found in significantly higher relative abundance in the CS compared with the VD infants at 1 month (p<0.0001; see online supplementary table S4). Following 6 months of age, there was a gradual increase in previous low abundant genera in both VD and CS infants. At 24 months of age, Bacteroides and several genera belonging to the Clostridia class, for example Ruminococcus, a dominant member of the adult microbiota, also dominated the infant microbiota.

Genus level microbiota composition in mothers and their infants at 1 week, 1, 3, 6, 12 and 24 months. The mean relative abundances (%) of the most abundant bacterial genera in the 15 vaginally delivered infants (A) and nine caesarean section infants (B) as well as in their mothers are shown. Only genera comprising ≥1% of the total community were included. Ac, Actinobacteria; Ba, Bacteroides; Fi, Firmicutes; Pr, Proteobacteria.
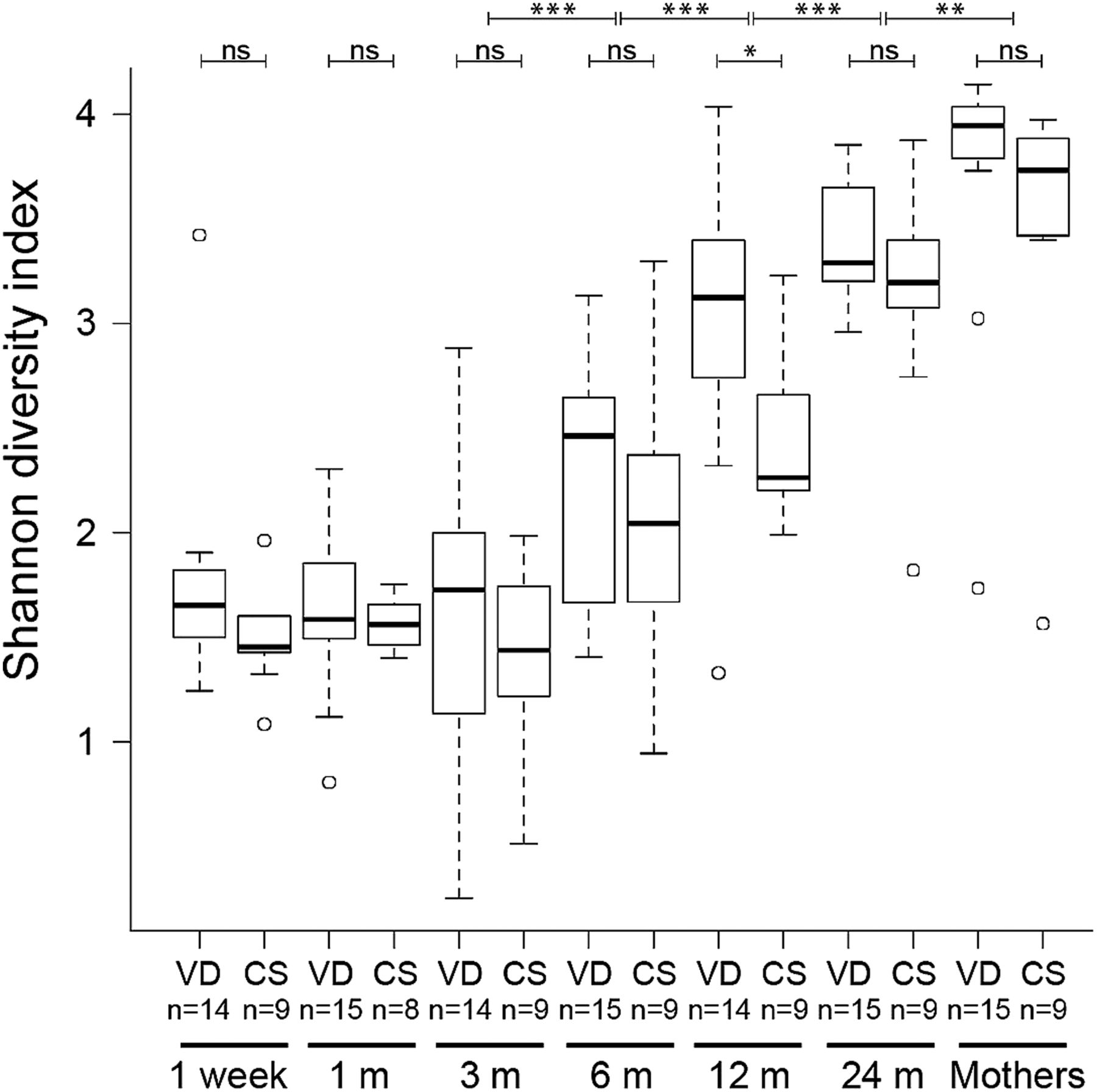
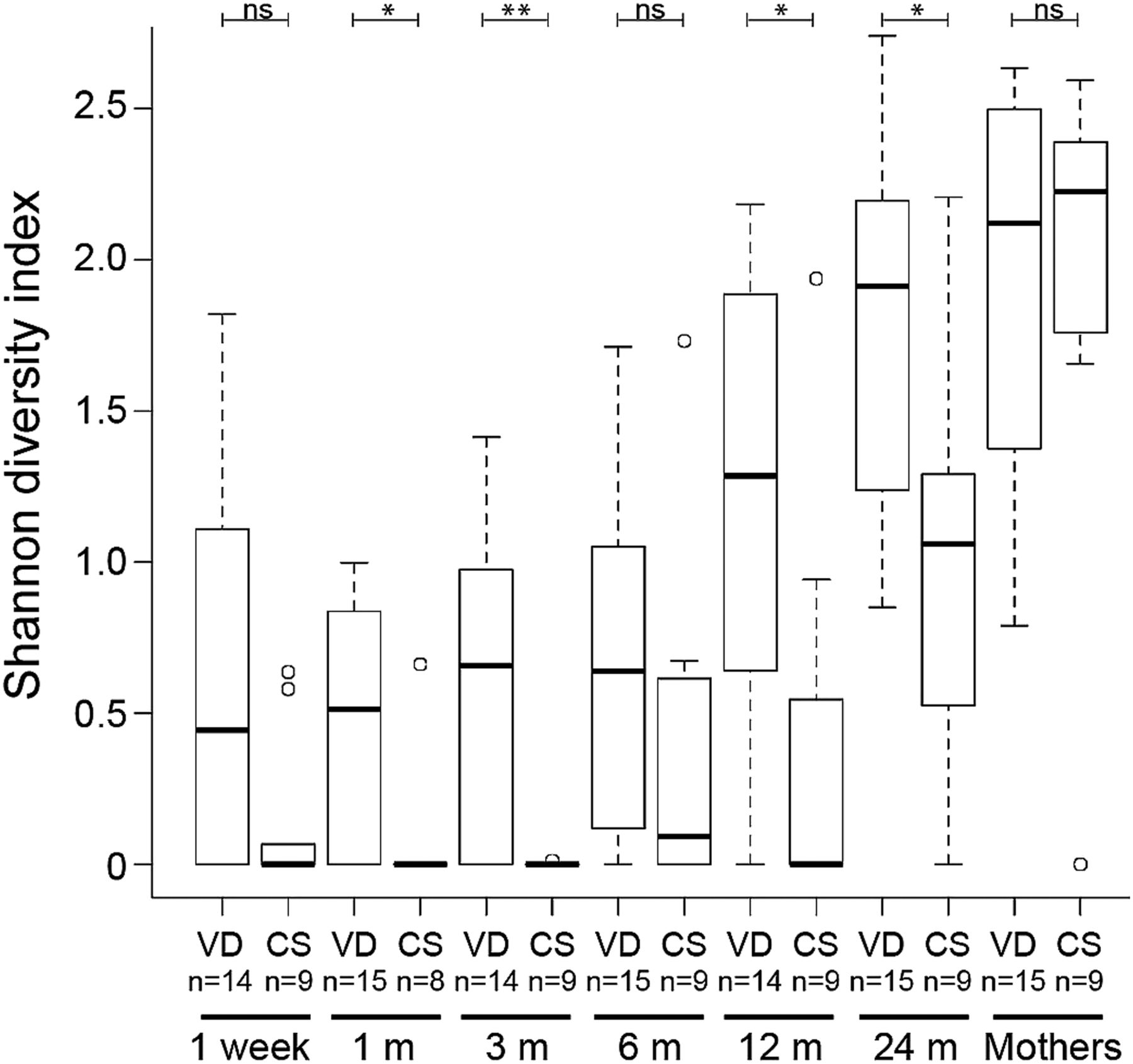
The gradual shift in community composition was accompanied by an increase in α-diversity over time, with a significant increase in Shannon diversity index between each pair of succeeding time points from 3 months of age onwards (figure 3). Similar results were obtained for evenness (Pielou's index; see online supplementary figure S1A) and estimated richness (Chao1; see online supplementary figure S1B). The low increase in diversity during the first 3 months may be related to that most (83%) infants were exclusively breast fed up to this age (see online supplementary table S1). CS delivery was associated with significantly lower total microbiota diversity when considering all time points in the infants (p=0.047 with repeated measures ANOVA; see online supplementary table S5). At individual time points, the total microbiota diversity was significantly lower in the CS delivered infants at 12 months (see online supplementary table S5; figure 3). The total microbiota diversity did not differ significantly between the VD and CS mothers (see online supplementary table S6). Narrowing the analysis to the phylum level, CS delivery was associated with a lower diversity of the Bacteroidetes phylum when considering all time points (p=0.002; see online supplementary table S5). For individual time points, the Bacteroidetes diversity was significantly lower in the CS infants than VD infants at 1, 3, 12 and 24 months (figure 4; see online supplementary table S6). The other phyla, Firmicutes, Proteobacteria and Actinobacteria, did not display any consistent differences in Shannon diversity between the groups, although diversity of Firmicutes was significantly lower at 12 months and Proteobacteria at 24 months in the CS infants (see online supplementary table S6).

Increase in faecal microbiota α-diversity over time. Distributions of Shannon diversity indices displayed for the 15 vaginally delivered (VD) infants and nine caesarean section (CS) infants at 1 week, 1, 3, 6, 12 and 24 months, and for their mothers. In all, 50% of the data points reside within boxes, 75% within whiskers and median values are indicated by horizontal lines within boxes (circles indicate individual values). Wilcoxon signed rank tests were conducted to compare Shannon diversity between adjacent time points, and Wilcoxon rank-sum tests to compare diversity between delivery modes within time points; ***p<0.001, **p<0.01 and *p<0.05.
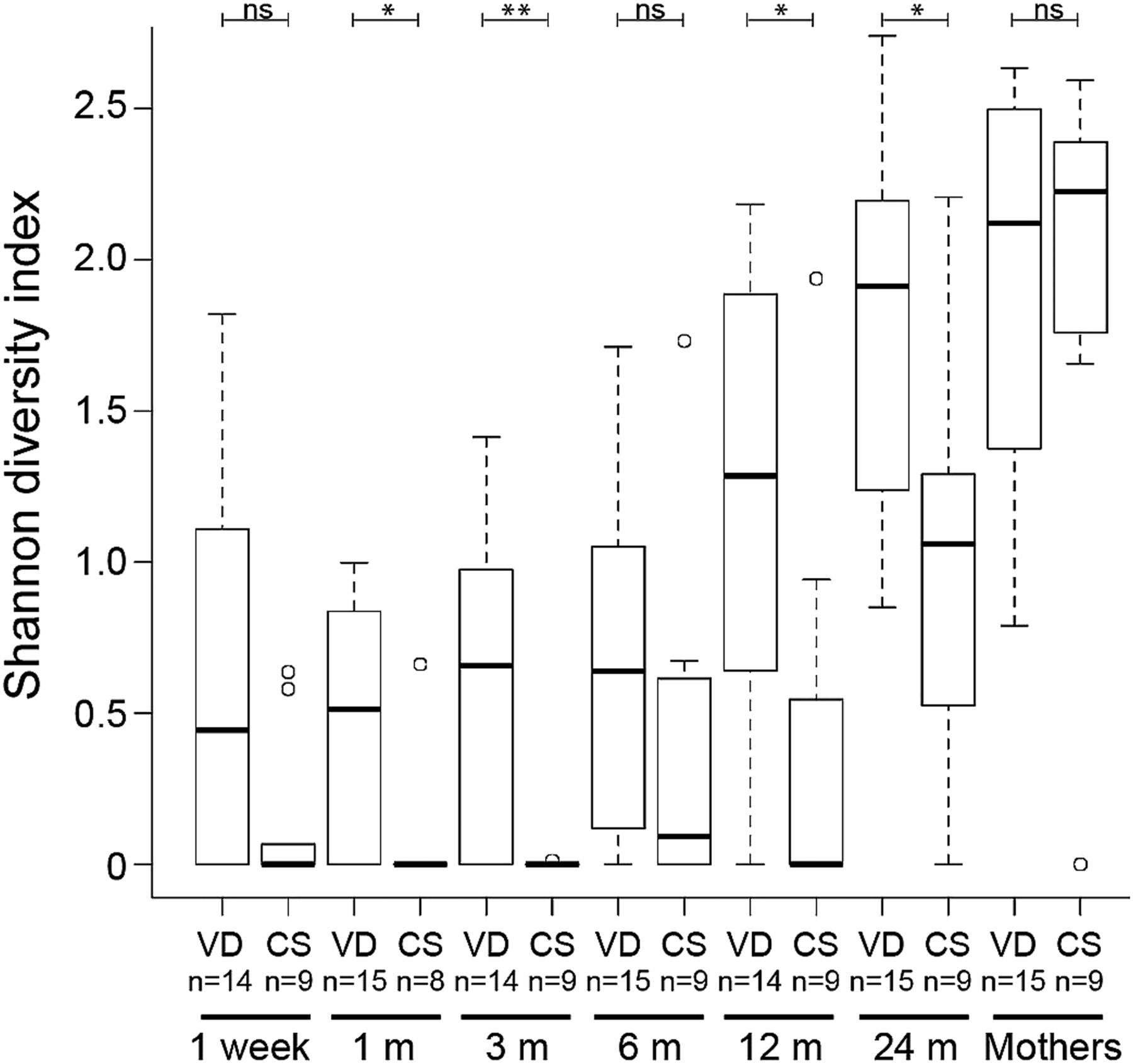
Increase in Bacteroidetes α-diversity over time. Distributions of Shannon diversity indices displayed for the 15 vaginally delivered (VD) infants and nine caesarean section (CS) infants at 1 week, 1, 3, 6, 12 and 24 months, and for their mothers. **p<0.01 and *p<0.05. In all, 50% of the data points reside within boxes, 75% within whiskers and median values are indicated by horizontal lines within boxes (circles indicate individual values).
A principal coordinates analysis plot based on pair-wise sample community differences calculated with UniFrac23 is illustrated in figure 5. As shown in a previous study31 the microbial communities became more uniform across infants over time, which is also evident when comparing the distributions of pair-wise community differences at each time point (see online supplementary figure S2). By 24 months of age, the communities closely resembled those of the mothers (figure 5). Although community composition converged to an adult-type microbiota, diversity estimates were still significantly lower at 24 months than in the mothers (figure 3; see online supplementary figure S1). Birth weight, antibiotic intake during the time course, sex and breast feeding had no apparent impact on the microbiota composition at any time point (see online supplementary figures S3–S6).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Individuality and convergence of infant microbiota. Principal coordinates analysis was performed on all pair-wise community differences (calculated with UniFrac23), and samples from infants at 1 week, 1, 3, 6, 12 and 24 months, and from the mothers are highlighted in the different boxes. Vaginally delivered and caesarean section infants/mothers are displayed in red and blue, respectively.
In order to investigate possible mother–child transmission of bacteria, the presence of specific unique sequences (sequence types) was compared between infants and their mothers, as well as between infants and other mothers. For each mother–child pair, we calculated the fraction of the sequence types found in the child that were also found in the mother (number of shared sequence types/number of child sequence types). The VD infants shared a significantly higher proportion of sequence types with their own mother than with the other mothers at 1 week and 24 months when considering all bacterial taxa (see online supplementary table S7). Considering one phylum at a time, Bacteroidetes displayed a significantly higher overlap at 1 and 6 months, and Firmicutes at 6 and 24 months. No significant overlap was observed for the CS delivered infants at any time point.
CS was associated with moderately lower levels of the Th1-associated chemokines CXCL10 and CXCL11 (table 1). The levels of the Th2-chemokines CCL17 and CCL22 did not differ significantly between the birth modes.
Repeated measures ANOVAs to test whether there were any significant differences in CXCL10 and CXCL11 levels during the first 2 years of life between caesarean section (CS) and vaginally delivered (VD) infants
Discussion
Microbial colonisation of the infant gut gastrointestinal tract is important for the postnatal development of the immune system. In this study, CS delivered infants who are not entering the birth canal of the mother either lacked or displayed a delayed colonisation of one of the major gut phylum, the Bacteroidetes. The colonisation of this phylum was delayed by up to 1 year for some infants. The total microbiota diversity was also lower in the CS infants, probably largely as a consequence of the lack of this phylum. This was not a consequence of antimicrobial treatment, as none of the CS mothers were given antibiotics before surgery and the microbial diversity did not differ between mothers who were given antibiotics prophylactically and those who did not receive antibiotics. Comparisons of intestinal microbiota have not conclusively confirmed bacterial transmission,31 ,32 although the genus Bacteroides has been proposed to be transmitted from the maternal gut.33 ,34 Our study corroborates earlier studies reporting a delayed colonisation of Bacteroides in babies delivered by CS.4 ,5 In addition, our study provides evidence that specific lineages of the intestinal microbiota, as defined by 16S rRNA gene sequences, are transmitted from mother to child during vaginal delivery.
It is important to note that bacterial composition changes as a consequence of freezing the faecal samples35 ,36 and that PCR amplification can induce taxon-specific biases. However, there were no significant differences in storage times between the VD and CS samples and since the same primer pair and PCR conditions were used for all samples, these effects should not contribute to the observed differences in microbiota composition and α-diversity between the sample groups.
The genus Enterococcus, which is a typical faecal bacterium, is usually acquired during the first week of life.37 Colonisation has previously not been shown to depend on delivery mode, suggesting other sources in addition to the maternal intestinal microbiota,5 such as the environment38 and breast milk.39 We found that CS infants had a higher relative abundance of Enterococcus at 1 month of age, suggesting that the lack of bacteria transmitted through vaginal delivery favours the growth and colonisation of enterococci.
Appropriate microbial stimulation during infancy is required for the development of a more balanced immune phenotype, including maturation of Th1-like responses and appropriate development of regulatory T cell responses.6 ,40 ,41 It is well known that early life events occurring during critical windows of immune development can have long-term impact on immune-mediated diseases such as allergy,15 ,42 ,43 diabetes and inflammatory bowel disease. We hypothesised that early differences in the gut microbiota could shape later immune responsiveness, influencing the Th1 maturation trajectory. Our findings of a lower microbial diversity in the CS infants and lower circulating levels of the Th1-related chemokines CXCL10 and CXCL11 support this view. Previous studies have shown that Bacteroides fragilis exert strong effects on the immune system. This is mediated by the capsular polysaccharide, which enhances T cell mediated immune responses and affects the Th1/Th2 balance.44 ,45 Furthermore, Bacteroides thetaiotamicron is also known to affect the immune system.46 Thus, the lower abundance of Bacteroides among the CS infants may be a contributing factor to the observed differences in Th1-associated chemokines. Future studies with larger sample sizes will be able to address the effects of individual microbes on chemokine levels.
With few exceptions,31 previous studies have reported Bifidobacterium to be one of the dominant genera of the early infant intestinal microbiota,4 ,33 ,47––49 and changes in relative abundance of this genus have been related to delivery mode.4 ,16 ,34 Also in the present study, Bifidobacterium was the dominant genus from 1 to 12 months of age with a gradual decline following weaning. The abundance was not affected by delivery mode, however, and we could not detect any significant overlap in the mothers’ and babies’ rRNA sequences. Hence, Bifidobacterium could primarily be transmitted from the breast milk, and to a lesser extent from the intestinal micobiota, as suggested but not confirmed previously.50
In accordance with recent studies,31 ,47 ,48 our results demonstrate considerable individual differences in the microbial succession during the first year of life. This is probably a result of differences in time of weaning, and incidental exposure of bacteria from the environment. Community composition converges to an adult-like state within 2 years. However, even at 2 years the microbiota appears not to be fully developed, since the diversity was significantly lower than in mothers. This was also evident in a recent study reporting a lower microbial diversity in a 2.5-year-old child than in its mother.51
An association between CS delivery and the development of allergic disease has been observed in several studies52 ,53 and a lower microbial diversity has been observed in allergic infants before onset of disease.13 We conclude that CS is associated with a lower bacterial diversity during the first 2 years of life, a lower abundance and diversity of the phylum Bacteroidetes, and lower circulating levels of Th1-associated chemokines during infancy.
Acknowledgments
We thank Mrs Lena Lindell, Linköping, Mrs Elisabeth Andersson, Norrköping, Mrs Linnea Andersson, Jönköping, and Mrs Eivor Folkesson, Motala, Dr Göran Oldaeus, Jönköping, and Dr Ted Jacobsson, Linköping, for their brilliant and enthusiastic work guiding the families through the study and all the sampling procedures. We also thank Ms Martina Abelius, Mrs Anne-Marie Fornander and Ms Anna Forsberg in Linköping for excellent technical assistance.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Files in this Data Supplement:
- Data supplement 1 - Online figure 1
- Data supplement 10 - Online table 4
- Data supplement 11 - Online table 5
- Data supplement 12 - Online table 6
- Data supplement 13 - Online table 7
- Data supplement 2 - Online figure 2
- Data supplement 3 - Online figure 3
- Data supplement 4 - Online figure 4
- Data supplement 5 - Online figure 5
- Data supplement 6 - Online figure 6
- Data supplement 7 - Online table 1
- Data supplement 8 - Online table 2
- Data supplement 9 - Online table 3
Press release
Files in this Data Supplement:
- Press release - Press release
Footnotes
-
Contributors Conception and design: HEJ, TRA, MCJ, BB, CJ, LE, AFA. Analysis and interpretation of data: HEJ, TRA, MCJ, KH, CQ, AFA. Drafting the article: HEJ, TRA, MCJ, BB, AFA. Final approval of submitted version: HEJ, TRA, MCJ, KH, CQ, CJ, BB, LE, AFA.
-
Funding This work was supported by the Ekhaga Foundation and the Söderbergs Foundation to LE and by the Swedish Research Council (K2011-56X-21854-01-06), the Research Council for the South-East Sweden, the Swedish Asthma and Allergy Association, the Olle Engkvist Foundation, the Vårdal Foundation–for Health Care Sciences and Allergy Research to MJ and by the Swedish Research Councils VR (621-2011-5689) and FORMAS (215-2009-1174) to AA. KH was funded through a direct grant from Unilever.
-
Competing interests None.
-
Ethics approval The human ethic committee at Linköping University, Linköping, Sweden.
-
Provenance and peer review Not commissioned; externally peer reviewed.