The Emerging Role of Vitamin C as a Treatment for Sepsis
by
 , , and
, , and
Markos G. Kashiouris
1,* ,
,
Michael L’Heureux
1,
Casey A. Cable
1,
Bernard J. Fisher
1,
Stefan W. Leichtle
2 and
Alpha A. Fowler
1 1
Department of Internal Medicine, Division of Pulmonary Disease and Critical Care Medicine, Virginia Commonwealth University School of Medicine, 1200 E Broad St., P.O. Box 980050, Richmond, VA 23298, USA
2
Department of Surgery, Division of Acute Care Surgical Services, Virginia Commonwealth University School of Medicine, 1200 E Broad St., P.O. Box 980454, Richmond, VA 23298, USA
*
Author to whom correspondence should be addressed.
Nutrients 2020, 12(2), 292; https://doi.org/10.3390/nu12020292
Submission received: 24 December 2019
/
Revised: 15 January 2020
/
Accepted: 16 January 2020
/
Published: 22 January 2020
(This article belongs to the Special Issue Vitamin C: From Bench to Bedside)
Abstract
:Sepsis, a life-threatening organ dysfunction due to a dysregulated host response to infection, is a leading cause of morbidity and mortality worldwide. Decades of research have failed to identify any specific therapeutic targets outside of antibiotics, infectious source elimination, and supportive care. More recently, vitamin C has emerged as a potential therapeutic agent to treat sepsis. Vitamin C has been shown to be deficient in septic patients and the administration of high dose intravenous as opposed to oral vitamin C leads to markedly improved and elevated serum levels. Its physiologic role in sepsis includes attenuating oxidative stress and inflammation, improving vasopressor synthesis, enhancing immune cell function, improving endovascular function, and epigenetic immunologic modifications. Multiple clinical trials have demonstrated the safety of vitamin C and two recent studies have shown promising data on mortality improvement. Currently, larger randomized controlled studies are underway to validate these findings. With further study, vitamin C may become standard of care for the treatment of sepsis, but given its safety profile, current treatment can be justified with compassionate use.
1. Introduction
Sepsis is a medical emergency that occurs due to a dysregulated host response to infection, resulting in life-threatening organ dysfunction [1]. The incidence of sepsis continues to rise in hospitals and intensive care units worldwide: approximately 31 million cases of sepsis occur globally every year, with approximately 6 million deaths [2]. In the United States, an estimated 535 cases of sepsis occur annually per 100,000 people, accounting for more than USD 23 billion in annual U.S. hospital expenditures in 2013 [2,3]. Despite new advances in critical care support techniques, 30–45% of patients die following hospitalization with severe sepsis and septic shock [4,5]. Sepsis can affect every organ system to cause morbidity and mortality. One particularly devastating complication of sepsis is acute respiratory distress syndrome (ARDS), a severe form of sepsis-induced lung injury. Compared with other forms of acute lung injury, patients with sepsis-induced lung injury spend more days on the mechanical ventilator and have a higher mortality rate [6]. Sepsis disproportionately affects elderly patients with comorbidities, such as impaired immune function and limited functional status [7]. The most common infectious source of sepsis in patients is pneumonia, followed by intra-abdominal and endovascular infections [8].
In sepsis, reactive oxygen and nitrogen species are generated, leading to the oxidant-induced activation of transcription factors that drive potent inflammatory cytokine and chemokine expression [9]. Oxidants produced during sepsis promote endothelial dysfunction, injuring surface glycocalyx proteins (e.g., syndecan-1) as well as damaging cell membranes and promoting the expression of intercellular adhesion molecules. During sepsis, endothelial cell damage leads to the loss of microvascular barrier function, resulting in enhanced vascular permeability. Furthermore, oxidant-induced expression of inflammatory cytokines and chemokines upregulates endothelial cell surface adhesion molecules, which leads to activated neutrophil and platelet adhesion in the microvasculature. Despite uncovering the complex multicellular activation pathways that drive tissue injury, a “cure” remains elusive. Clinical trials have included over 15,000 patients and spent over one billion US dollars in randomized clinical trial costs [10]. These trials have targeted mediators of inflammation or coagulation such as statin agents [11,12], activated protein C [13], and monoclonal antibody to endotoxin [14], but have not safely reduced sepsis mortality, suggesting that single-target therapy fails to meet the challenges of treating the complex pathophysiology of sepsis. Overall, this accumulated knowledge strongly suggests that a more “pleotropic” (i.e., broad-based) form of therapy that can interrupt multiple pathways is required to uncover the “holy grail” of treating sepsis. At present, in 21st-century critical care practice, antibiotics, “source control,” and maintaining hemodynamic stability with fluid administration and vasopressors constitutes the mainstay therapy for sepsis. More recently, there has been emerging evidence for the use of vitamin C as a treatment of sepsis.
2. Pharmacology and Physiology of Vitamin C
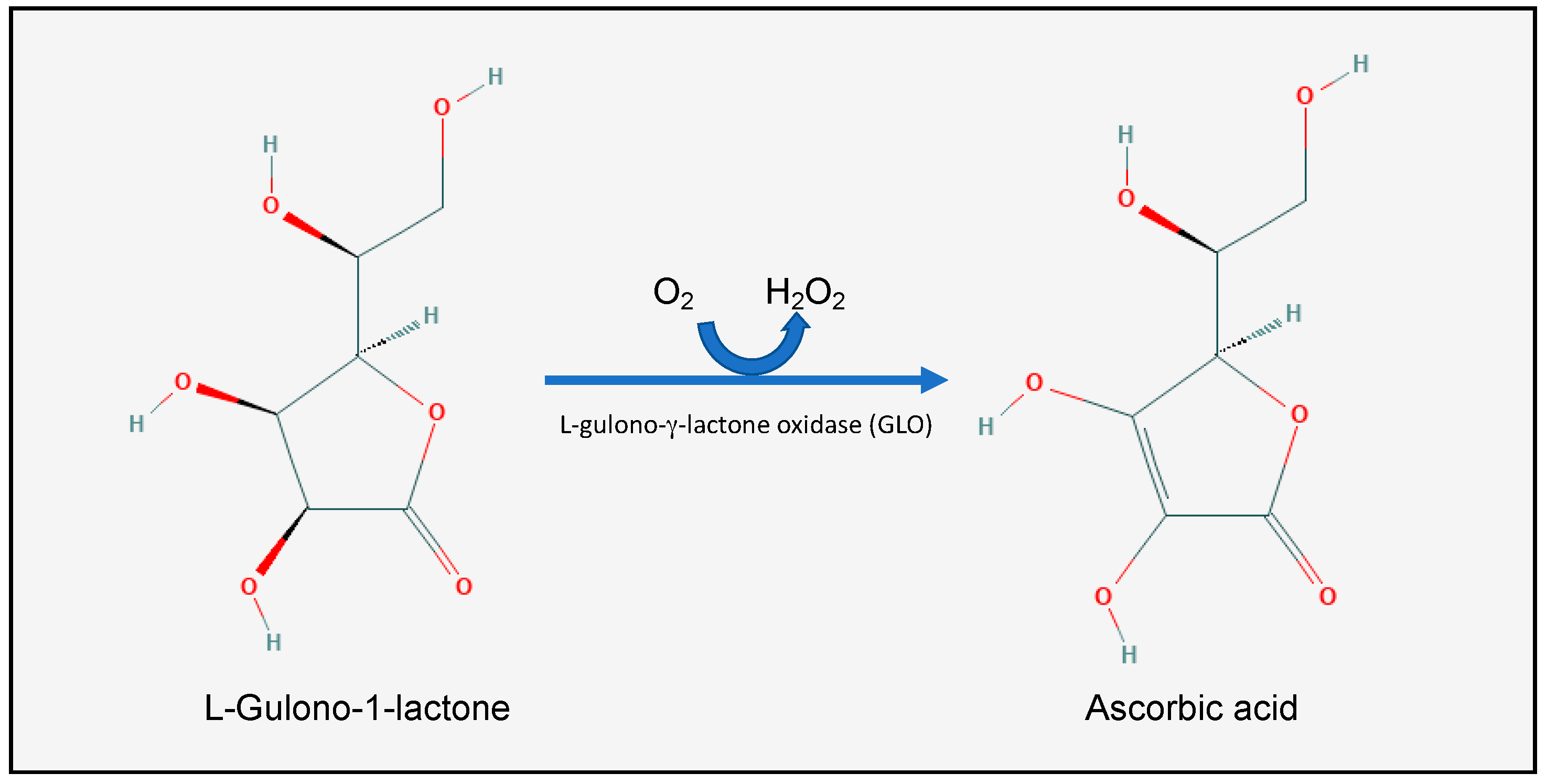
L-Ascorbic acid (AA), or vitamin C (Figure 1), is a water-soluble vitamin which is found in all fruits and vegetables, but is particularly concentrated in citrus fruits, green peppers, strawberries, broccoli, green leaves, and potatoes [15]. It was first isolated between 1927 and 1930 from ox adrenals by the Nobel laureate Albert Szent-Györgyi at Cambridge University and the Mayo Clinic. Most animal species synthesize vitamin C in the kidneys or the liver. Humans, some mammals (such as gorillas, monkeys, bats, and guinea pigs), birds, and fish, however, have lost the ability to synthesize vitamin C [16]. More specifically, these species lost the ability to synthesize the l-gulono-γ-lactone oxidase (GLO) enzyme, which catalyzes the last step in the vitamin C synthesis: oxidizing l-Gulono-1-lactone into l-Ascorbic acid [16]. The human GLO gene was inactivated by random mutations somewhere between 38 and 92 million years ago [17]. This gene loss was compatible with survival, however, because of the abundance of vitamin C in the human diet [18].
2.1. Vitamin C Homeostasis
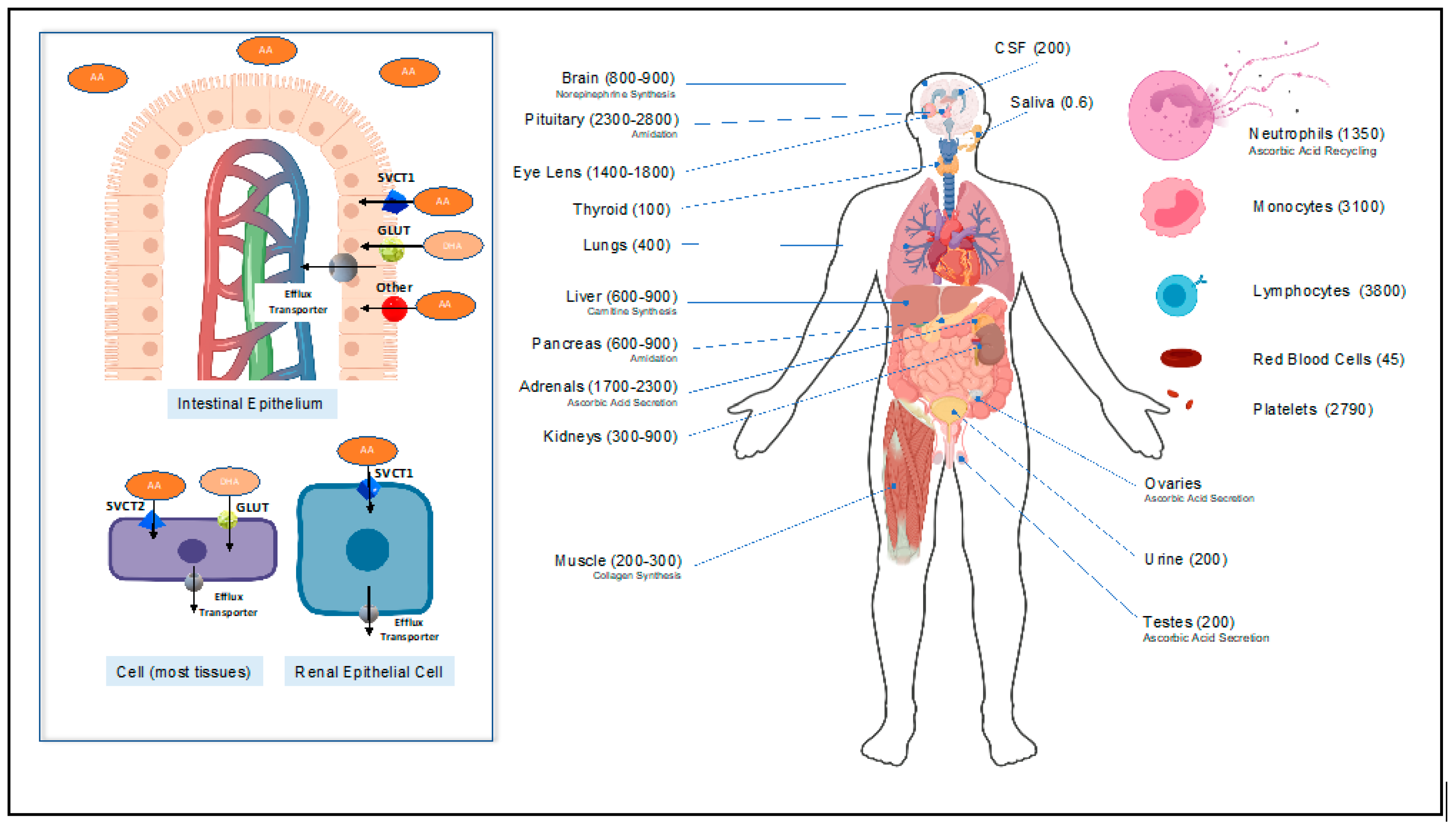
Since humans do not synthesize vitamin C, they depend on vitamin C dietary intake to stay alive. The human body stores approximately 1.5 g of vitamin C, and scurvy develops when the stores drop below 0.3 g, which is equivalent to a plasma concentration of less than 11 uM [19,20]. Normal plasma vitamin C concentrations range from 50 to 80 uM, but tissue concentrations can often reach millimolar levels (with the exception of saliva) [19]. Vitamin C is moved intracellularly by carrier proteins known as sodium-dependent vitamin C transporters (SVCT). Two SVCT isoforms exist—SVCT1 and SVCT 2—and these isoforms are highly conserved glycoproteins with 12 transmembrane domains [21,22]. While almost all body tissues (including white blood cells and platelets) express SVCT2, SVCT1 is expressed exclusively by intestinal epithelial cells, the proximal convoluted renal tubules, and the liver [21,22].
When taken orally, SVCT1 in the apical membrane of the small intestinal epithelial lumen (Figure 2) actively transports vitamin C into the epithelial cells [21]. Additionally, in the intestinal lumen, an oxidized form of vitamin C (dehydroascorbic acid (DHA)), is transported by the abundant facilitative glucose transporters GLUT into the intestinal epithelium [21]. More specifically, GLUT2 and GLUT8, which are expressed on the apical (luminal) membrane of the intestinal epithelium, are integral to DHA uptake [23]. Following absorption, the water-soluble vitamin C molecule distributes in the blood and the extracellular compartment. Tissues then take up the vitamin C via the SVCT2 transporters (Figure 2) [23]. Red blood cells are an exception, as they take up DHA and reduce it via the glutaredoxin protein or glutathione [24]. Finally, the kidneys freely filter vitamin C and reabsorb it via SVCT1 in the proximal convoluted renal tubules [23].
Pathologically, vitamin C deficiency presents as scurvy, but many disease states alter vitamin C homeostasis. Vitamin C concentrations are often low in acute illnesses such as myocardial infarction, acute pancreatitis, sepsis, and critical illness in general [26,27,28]. Moreover, aging humans (who have a higher baseline critical illness mortality) require more dietary vitamin C to reach a desired plasma ascorbate concentration. These observations are confirmed in animal studies that show diminished SVCT1 mRNA expression with aging, resulting in a decline in the capacity of cells to absorb vitamin C [28,29].
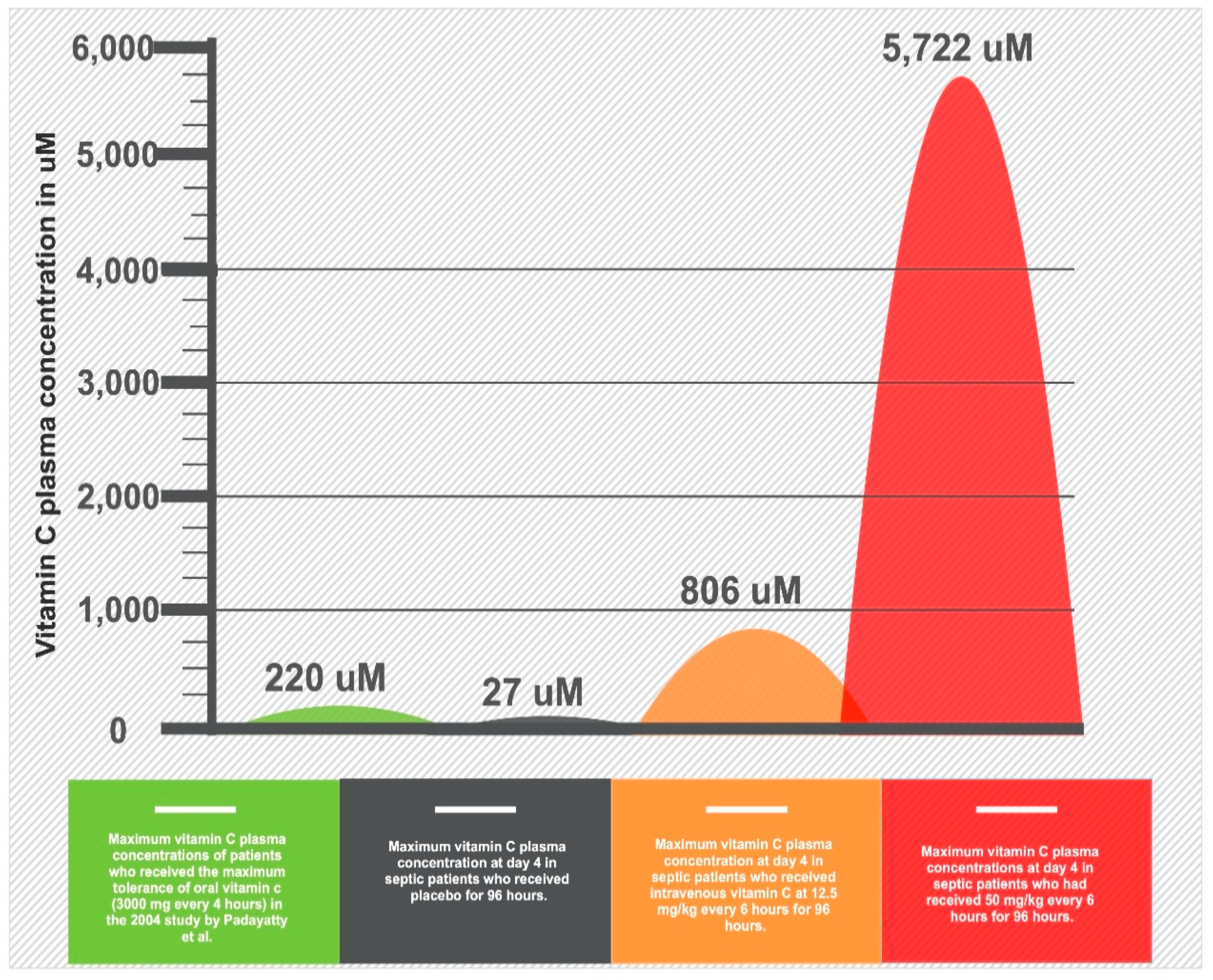
The pharmacokinetics of vitamin C absorption were explored in a landmark study of human volunteers [30]. The bioavailability, which is the percentage (%) of the drug that reaches the systemic circulation, is 100% for a single oral dose of 200 mg but decreases to 33% with a single dose of 1250 mg [30]. This is because the intestinal SVCT1 transporter achieves maximal saturation around 500–1000 mg (Figure 3). In addition, the bioavailability does not decrease linearly because of alterations in renal vitamin C excretion [30]. Overall, the maximum peak plasma concentration that was achieved with long-term, high-dose oral dosing (i.e., 3 g every 4 h) is 220 uM [31]. Intravenous vitamin C administration, however, can bypass the limitations of SVCT1-induced bioavailability to achieve 70-fold higher plasma concentrations compared to oral administration [31]. In contrast to oral intake, intravenous administration can achieve a peak plasma concentration up to 15,000 uM [31]. It is key to understand that most of the earlier trials did not appreciate the differential pharmacokinetics of vitamin C and made assumptions that the plasma levels achieved with oral administration are equivalent with the intravenous administration.
2.2. Pleiotropic Physiologic Functions of Vitamin C
Vitamin C has numerable physiologic molecular functions due to its role as an electron donor/reducing agent [25]. These include direct antioxidant properties and cofactor properties for a wide array of enzymes in a wide variety of cellular structures and organelles, resulting in varied tissue and organ-system effects (Table 1).
2.3. Vitamin C’s Mechanism of Action in Sepsis and ARDS
Subnormal plasma vitamin C concentrations are common in critically ill patients and, in particular, patients with sepsis [26,27,32,113]. Furthermore, lower vitamin C levels correlate with higher incidence of organ failure and worse outcomes in septic patients [93]. In fact, very low plasma vitamin C levels, averaging around 18 uM, are a predictable feature in severely septic patients [114]. This is caused by the explosive cytokine release present in sepsis, which interferes with the cellular regulation of vitamin C absorption. Seno et al. showed that inflammatory cytokines, such as TNFα and IL-1β, known to be present in sepsis, negatively regulate endothelial SVCT2 activity; this results in the depletion of intracellular vitamin C levels [115]. In addition, during this overwhelming disease state of oxidative stress and increased reactive oxygen species (ROS) production, there is increased vitamin C consumption by the somatic cells and by leukocyte turnover [40]. As Figure 2 suggests, leukocytes can have an up to 100-fold increase in the concentration of vitamin C compared to plasma. The increased production and turnover of those cells contributes to vitamin C depletion in sepsis [40].
Many of the physiologic roles of vitamin C are important in patients with sepsis. These include the key antioxidant properties of vitamin C, scavenging reactive oxygen species, repletion of other crucial body antioxidants vitamin E and glutathione [32,35,38,39,42,49,62,64,66,68,116], and cardiovascular benefits by supporting endogenous norepinephrine, dopamine, and vasopressin production [41,117,118]. Furthermore, vitamin C protects against the loss of epithelial and endothelial barriers and enhances neutrophil function in a multidimensional way. Moreover, vitamin C promotes lymphocytic and neutrophilic activity while attenuating neutrophil necrosis and NETosis (neutrophil extracellular trap), which contributes to multiorgan failure [69,70,71,72,73,74,75,76,77,78,79,80,81,82,83,119,120,121,122]. Vitamin C also regulates nuclear cellular responses to stress and hypoxia by regulating HIF-1α [40,49,52,62,123], produces NF-κB epigenetic modifications [124] through its ability to de-methylate histones [75,82,87,88,89,125,126], regulates pro-inflammatory and coagulation gene expression [28,41,42,44,123,125,126,127], and orchestrates the immune system and circulating cytokine homeostasis in pleotropic ways (Table 1). The combination of vitamin C’s vital functions and its depletion in septic states justifies the use of high-dose intravenous vitamin C (HDIVC) in the early phases of severe sepsis and septic shock [116].
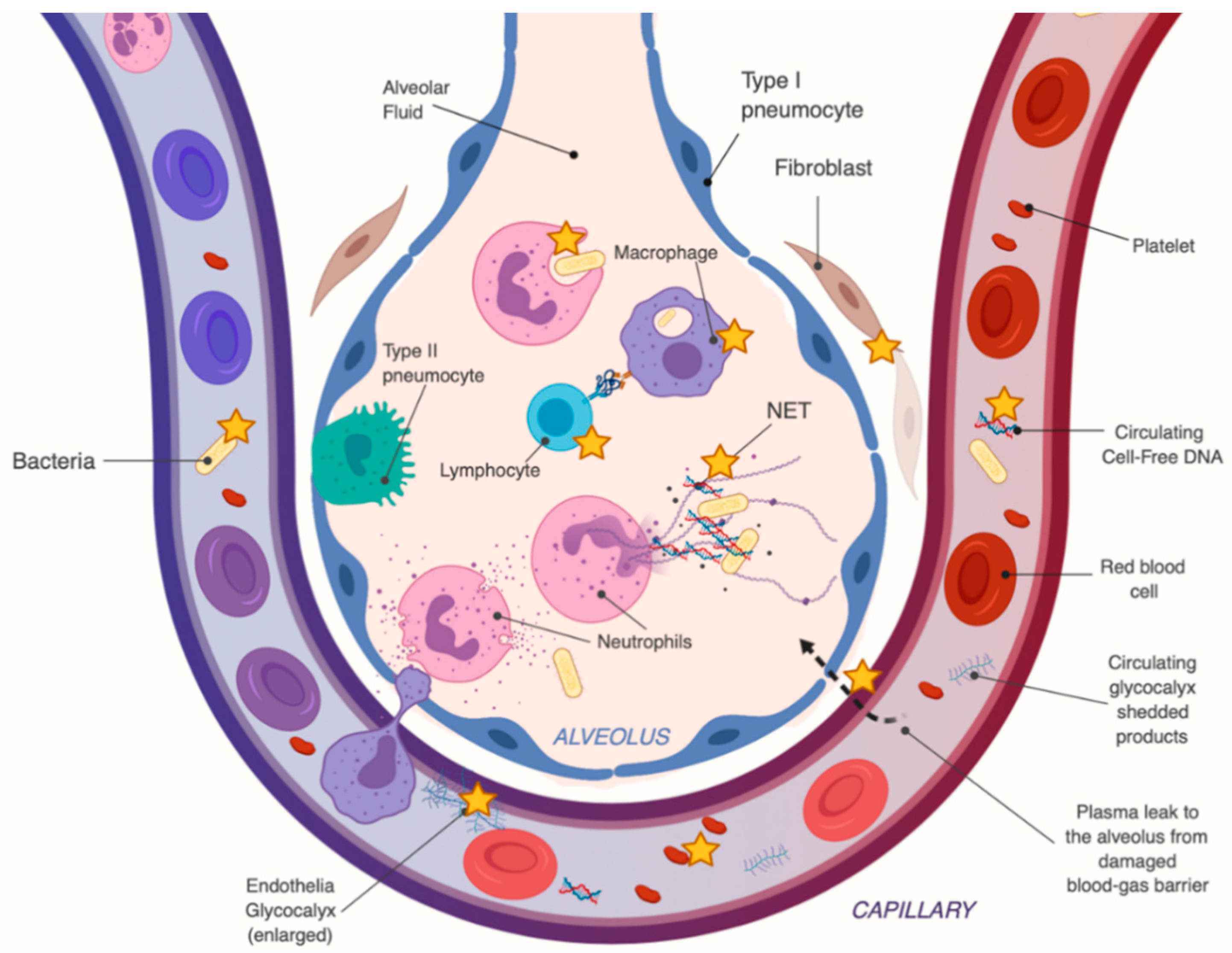
Vitamin C also has effects in the septic patient that are more specific to sepsis-induced ARDS, which are summarized in Figure 4. These include enhanced lung epithelial barrier function (i.e., via claudins and occludins) and by epigenetic and transcriptional enhancement of protein-channels which regulate the alveolar fluid clearance, such as aquaporin-5, cystic fibrosis transmembrane regular (CFTR), epithelial sodium channels (ENaC) and Na+/ K+ ATPases [128,129]. There is rising evidence that HDIVC treatment in sepsis-induced ARDS results in significantly lower levels of circulating cell-free DNA [108], which have been associated with multiorgan failure. HDIVC treatment also resulted in significant reduction of plasma circulating syndecan-1, a component of the endothelial glycocalyx, whose levels closely correlate with and predict mortality in patients with severe sepsis and ARDS [107].
3. Clinical Trials
The initial data on the clinical use of vitamin C was obtained in animal models. Subsequently, several completed clinical trials contributed evidence for the therapeutic effects of HDIVC in human sepsis. The first study, published in 1986, treated 16 ARDS patients with intravenous vitamin C (1000 mg IV every 6 h) plus antioxidants (N-acetylcysteine, selenium, and vitamin E) versus 16 ARDS patients who received the standard care at that time (i.e., control group) [130]. There was a dramatic reduction in mortality in the vitamin C group—37% versus 71% in the standard care group (p < 0.01) [130]. A phase I trial in 2014 [114] proved that plasma vitamin C levels in patients with severe sepsis were low, almost at scorbutic levels, and that HDIVC administration had a dose-dependent effect in the prevention of multi-organ failure, as measured by the Sequential Organ Failure Assessment (SOFA) scores [131]. Patients who received a total of 200 mg/kg/day of HDIVC for 4 days (administered in 50 mg/kg/dose, every 6 h), had significantly lower SOFA scores than placebo, and even lower scores than the patients who received lower-doses of IV vitamin C (50 mg/kg/day administered at 12.5 mg/kg/dose, every 6 h for 4 days). In this trial, the patients in the HDIVC group (200 mg/kg/day) achieved plasma levels of up to 3000 uM at day 4. The patients receiving HDIVC also demonstrated statistically lower inflammatory biomarker levels (C-Reactive protein and procalcitonin) and lower thrombomodulin levels, which is a marker of endothelial injury [114].
In 2016, a retrospective before–after study of 94 patients with severe sepsis and septic shock [132] compared patients who received hydrocortisone (50 mg IV every 6 h for 7 days or until ICU discharge), thiamine (200 mg IV every 12 h for 4 days or until ICU discharge) and HDIVC (6000 mg/day, in 4 divided doses for 4 days or until ICU discharge) to control. This study showed a 31.9% decrease in absolute hospital mortality between cases who received the triple-therapy and controls (8.5% vs. 40.4% respectively). A small randomized controlled trial, performed around the same time, of 28 patients with septic shock who received moderate doses of IV vitamin C (25 mg/kg every 6 h for 3 days) showed significantly lower mortality in patients who received IV vitamin C—14.3% vs. 64.3% [117]. The same trial found a significant reduction in average norepinephrine doses, total norepinephrine doses and total duration of norepinephrine infusion [117]. A subsequent meta-analysis of the three above studies found a significant benefit of intravenous vitamin C, with “marked reduction” in mortality and duration of vasopressor administration [133].
The largest trial completed on vitamin C to date, the CITRIS-ALI trial, was published in 2019 [134]. This multicenter, randomized, double-blinded trial included 167 patients with sepsis and ARDS who were randomized to receive 50 mg/kg every 6 h of HDIVC for 4 days versus placebo and showed statistically significant difference in 28-day all-cause mortality. The 28-day mortality was 29.8% in the vitamin C group versus 46.3% in the placebo group, although this was a secondary outcome. The statistical effect on mortality remained for up to 60 days following trial completion. The most dramatic reduction in mortality was noted during the period of HDIVC infusion (Figure 5). Furthermore, the HDIVC group had a strong trend towards more ventilator-free days (13.1 in the HDIVC group vs 10.6 in the placebo group mean difference, 2.47, 95% CI −0.90–5.85, p = 0.15), ICU-free days to day 28 (10.7 in HDIVC group vs. 7.7, in the placebo group, p = 0.03), and more hospital-free days (22.6 in HDIVC group vs. 15.5, respectively, p = 0.04). This trial did not find significant reductions in the SOFA scores, C-reactive protein, thrombomodulin or procalcitonin. Those biomarkers and scores, however, were not measured among the patients who “graduated” early from the ICU (a group that was heavily shifted towards the HDIVC group) or in those patients who died (heavily shifted towards the placebo group), indicating a strong selection bias, which makes these results difficult to interpret. Several other randomized controlled trials of HDIVC are under way, such as the VICTAS trial, and the Clinical Trials Network for the Prevention and Early Treatment of Acute Lung Injury (PETAL Network) is currently planning a randomized controlled trial of HDIVC for the prevention of ARDS.
4. Adverse Effects of Vitamin C Therapy
In all the sepsis trials mentioned above, HDIVC was found to be safe and no significant side-effects were identified. Additionally, two studies in non-medical patients did not report adverse side effects. The first was a study of infused vitamin C, 1000 mg every 8 h, combined with oral vitamin E for 28 days in 594 surgically critically ill patients and found a significantly lower incidence of acute lung injury and multiorgan failure, with no side effects [35]. The second was a study of infused vitamin C continuously at 66 mg/kg per hour for the first 24 h in patients with greater than 50% surface area burns showed that the therapy was well-tolerated with no reported side effects [135]. One proposed side effect of HDIVC is an increased propensity for oxalate kidney stone production, but this has not been shown in any clinical trials to date.
One consideration in utilizing vitamin C is that it is thought to cause an artefactual rise in point-of-care blood glucose readings by nearly all point-of-care devices [136,137]. It does not, however, raise blood glucose readings from a basic metabolic panel or glucose results using blood gas laboratories that employ hexokinase technology for analysis. This finding has recently been questioned by a review of five patients where the artifact was not appreciated [138]. For now, care must be taken to assure an accurate blood glucose level from a metabolic laboratory (i.e., basic metabolic panel (BMP)) or arterial blood gas panel (ABG) before initiating any insulin therapy, given the risk of hypoglycemia due to incorrect dosage of insulin from artefactual glucometer readings.
5. Conclusions
A plethora of laboratory, animal, and clinical studies are building a compelling case for a crucial role of HDIVC in the treatment of sepsis. Given the multitude of mechanisms of action, vitamin C may succeed where other possible sepsis treatments have previously failed, or facilitate the success of a multi-modal approach. Not all vitamin C treatments, however, are created equal. Because of limitations in bioavailability, oral administration does not allow for the therapeutic plasma levels required in critical conditions such as sepsis, septic shock, and ARDS. Additionally, we intentionally used the acronym HDIVC to highlight that is an entirely different therapy than oral vitamin C, or low-dose intravenous administration.
Many HDIVC randomized trials are under way and at the time of publication HDIVC use in clinical practice can be used compassionately, given that it is safe, but is not yet supported by the sepsis guidelines. Clinicians should carefully appraise the existing literature, understand the pharmacokinetics, physiology and clinical evidence of HDIVC in sepsis and other syndromes, and weigh the risks and benefits of vitamin C infusion jointly with the patient and/or the patient’s family.
Author Contributions
Conceptualization, A.A.F. and M.G.K.; writing—original draft preparation, M.G.K., S.W.L.; writing—review and editing, M.G.K., C.A.C., M.L., S.W.L., and B.J.F.; table and figure creation—M.G.K., M.L., B.J.F., S.W.L., and C.A.C. All authors have read and agree to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The primary authors (M.G.K., B.J.F., A.A.F.) were investigators on the CITRUS-ALI study.
References
- Singer, M.; Deutschman, C.S.; Seymour, C.W.; Shankar-Hari, M.; Annane, D.; Bauer, M.; Bellomo, R.; Bernard, G.R.; Chiche, J.-D.; Coopersmith, C.M.; et al. The Third International Consensus Definitions for Sepsis and Septic Shock (Sepsis-3). JAMA 2016, 315, 801–810. [Google Scholar] [CrossRef]
- Fleischmann, C.; Scherag, A.; Adhikari, N.K.J.; Hartog, C.S.; Tsaganos, T.; Schlattmann, P.; Angus, D.C.; Reinhart, K. International Forum of Acute Care Trialists Assessment of Global Incidence and Mortality of Hospital-treated Sepsis. Current Estimates and Limitations. Am. J. Respir. Crit. Care Med. 2016, 193, 259–272. [Google Scholar] [CrossRef] [PubMed]
- Walkey, A.J.; Lagu, T.; Lindenauer, P.K. Trends in sepsis and infection sources in the United States. A population-based study. Ann. Am. Thorac. Soc. 2015, 12, 216–220. [Google Scholar] [CrossRef] [PubMed]
- Andreu Ballester, J.C.; Ballester, F.; González Sánchez, A.; Almela Quilis, A.; Colomer Rubio, E.; Peñarroja Otero, C. Epidemiology of sepsis in the Valencian Community (Spain), 1995–2004. Infect. Control Hosp. Epidemiol. 2008, 29, 630–634. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J.; Vincent, J.-L.; Adhikari, N.K.J.; Machado, F.R.; Angus, D.C.; Calandra, T.; Jaton, K.; Giulieri, S.; Delaloye, J.; Opal, S.; et al. Sepsis: A roadmap for future research. Lancet Infect. Dis. 2015, 15, 581–614. [Google Scholar] [CrossRef]
- Sheu, C.-C.; Gong, M.N.; Zhai, R.; Chen, F.; Bajwa, E.K.; Clardy, P.F.; Gallagher, D.C.; Thompson, B.T.; Christiani, D.C. Clinical characteristics and outcomes of sepsis-related vs non-sepsis-related ARDS. Chest 2010, 138, 559–567. [Google Scholar] [CrossRef] [Green Version]
- Kaukonen, K.-M.; Bailey, M.; Suzuki, S.; Pilcher, D.; Bellomo, R. Mortality related to severe sepsis and septic shock among critically ill patients in Australia and New Zealand, 2000–2012. JAMA 2014, 311, 1308–1316. [Google Scholar] [CrossRef]
- Rhee, C.; Jones, T.M.; Hamad, Y.; Pande, A.; Varon, J.; O’Brien, C.; Anderson, D.J.; Warren, D.K.; Dantes, R.B.; Epstein, L.; et al. Centers for Disease Control and Prevention (CDC) Prevention Epicenters Program Prevalence, Underlying Causes, and Preventability of Sepsis-Associated Mortality in US Acute Care Hospitals. JAMA Netw. Open 2019, 2, e187571. [Google Scholar] [CrossRef] [Green Version]
- Salvemini, D.; Cuzzocrea, S. Oxidative stress in septic shock and disseminated intravascular coagulation. Free Radic. Biol. Med. 2002, 33, 1173–1185. [Google Scholar] [CrossRef]
- Marshall, J.C. Clinical trials of mediator-directed therapy in sepsis: What have we learned? Intensive Care Med. 2000, 26 (Suppl. 1), S75–S83. [Google Scholar] [CrossRef]
- Kruger, P.; Bailey, M.; Bellomo, R.; Cooper, D.J.; Harward, M.; Higgins, A.; Howe, B.; Jones, D.; Joyce, C.; Kostner, K.; et al. ANZ-STATInS Investigators–ANZICS Clinical Trials Group A multicenter randomized trial of atorvastatin therapy in intensive care patients with severe sepsis. Am. J. Respir. Crit. Care Med. 2013, 187, 743–750. [Google Scholar] [CrossRef] [PubMed]
- Truwit, J.D.; Bernard, G.R.; Steingrub, J.; Matthay, M.A.; Liu, K.D.; Albertson, T.E.; Brower, R.G.; Shanholtz, C.; Rock, P.; Douglas, I.S.; et al. Rosuvastatin for sepsis-associated acute respiratory distress syndrome. N. Engl. J. Med. 2014, 370, 2191–2200. [Google Scholar] [PubMed] [Green Version]
- Ranieri, V.M.; Thompson, B.T.; Barie, P.S.; Dhainaut, J.-F.; Douglas, I.S.; Finfer, S.; Gårdlund, B.; Marshall, J.C.; Rhodes, A.; Artigas, A.; et al. PROWESS-SHOCK Study Group Drotrecogin alfa (activated) in adults with septic shock. N. Engl. J. Med. 2012, 366, 2055–2064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziegler, E.J.; Fisher, C.J.; Sprung, C.L.; Straube, R.C.; Sadoff, J.C.; Foulke, G.E.; Wortel, C.H.; Fink, M.P.; Dellinger, R.P.; Teng, N.N. Treatment of gram-negative bacteremia and septic shock with HA-1A human monoclonal antibody against endotoxin. A randomized, double-blind, placebo-controlled trial. The HA-1A Sepsis Study Group. N. Engl. J. Med. 1991, 324, 429–436. [Google Scholar] [CrossRef] [PubMed]
- National Cancer Institute Ascorbic Acid (Code C285). Available online: https://ncit.nci.nih.gov/ncitbrowser/ConceptReport.jsp?dictionary=NCI_Thesaurus&ns=NCI_Thesaurus&code=C285 (accessed on 11 September 2019).
- Drouin, G.; Godin, J.-R.; Pagé, B. The genetics of vitamin C loss in vertebrates. Curr. Genom. 2011, 12, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Lachapelle, M.Y.; Drouin, G. Inactivation dates of the human and guinea pig vitamin C genes. Genetica 2011, 139, 199–207. [Google Scholar] [CrossRef]
- Pauling, L. Evolution and the need for ascorbic acid. Proc. Natl. Acad. Sci. USA 1970, 67, 1643–1648. [Google Scholar] [CrossRef] [Green Version]
- Granger, M.; Eck, P. Dietary vitamin C in human health. Adv. Food Nutr. Res. 2018, 83, 281–310. [Google Scholar]
- Hodges, R.E.; Hood, J.; Canham, J.E.; Sauberlich, H.E.; Baker, E.M. Clinical manifestations of ascorbic acid deficiency in man. Am. J. Clin. Nutr. 1971, 24, 432–443. [Google Scholar] [CrossRef] [Green Version]
- Tsukaguchi, H.; Tokui, T.; Mackenzie, B.; Berger, U.V.; Chen, X.Z.; Wang, Y.; Brubaker, R.F.; Hediger, M.A. A family of mammalian Na+-dependent L-ascorbic acid transporters. Nature 1999, 399, 70–75. [Google Scholar] [CrossRef]
- Daruwala, R.; Song, J.; Koh, W.S.; Rumsey, S.C.; Levine, M. Cloning and functional characterization of the human sodium-dependent vitamin C transporters hSVCT1 and hSVCT2. FEBS Lett. 1999, 460, 480–484. [Google Scholar] [CrossRef] [Green Version]
- Corpe, C.P.; Eck, P.; Wang, J.; Al-Hasani, H.; Levine, M. Intestinal dehydroascorbic acid (DHA) transport mediated by the facilitative sugar transporters, GLUT2 and GLUT8. J. Biol. Chem. 2013, 288, 9092–9101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuhn, V.; Diederich, L.; Keller, T.C.S.; Kramer, C.M.; Lückstädt, W.; Panknin, C.; Suvorava, T.; Isakson, B.E.; Kelm, M.; Cortese-Krott, M.M.; et al. Red blood cell function and dysfunction: Redox regulation, nitric oxide metabolism, anemia. Antioxid. Redox Signal. 2017, 26, 718–742. [Google Scholar] [CrossRef] [PubMed]
- Padayatty, S.J.; Levine, M. Vitamin C: The known and the unknown and Goldilocks. Oral Dis. 2016, 22, 463–493. [Google Scholar] [CrossRef] [Green Version]
- Schorah, C.J.; Downing, C.; Piripitsi, A.; Gallivan, L.; Al-Hazaa, A.H.; Sanderson, M.J.; Bodenham, A. Total vitamin C, ascorbic acid, and dehydroascorbic acid concentrations in plasma of critically ill patients. Am. J. Clin. Nutr. 1996, 63, 760–765. [Google Scholar] [CrossRef]
- Long, C.L.; Maull, K.I.; Krishnan, R.S.; Laws, H.L.; Geiger, J.W.; Borghesi, L.; Franks, W.; Lawson, T.C.; E Sauberlich, H. Ascorbic acid dynamics in the seriously ill and injured. J. Surg. Res. 2003, 109, 144–148. [Google Scholar] [CrossRef]
- Metnitz, P.G.; Bartens, C.; Fischer, M.; Fridrich, P.; Steltzer, H.; Druml, W. Antioxidant status in patients with acute respiratory distress syndrome. Intensive Care Med. 1999, 25, 180–185. [Google Scholar] [CrossRef]
- Fulzele, S.; Chothe, P.; Sangani, R.; Chutkan, N.; Hamrick, M.; Bhattacharyya, M.; Prasad, P.B.; Zakhary, I.; Bowser, M.; Ganapathy, V.; et al. Sodium-dependent vitamin C transporter SVCT2: Expression and function in bone marrow stromal cells and in osteogenesis. Stem Cell Res. 2013, 10, 36–47. [Google Scholar] [CrossRef] [Green Version]
- Levine, M.; Conry-Cantilena, C.; Wang, Y.; Welch, R.W.; Washko, P.W.; Dhariwal, K.R.; Park, J.B.; Lazarev, A.; Graumlich, J.F.; King, J.; et al. pharmacokinetics in healthy volunteers: Evidence for a recommended dietary allowance. Proc. Natl. Acad. Sci. USA 1996, 93, 3704–3709. [Google Scholar] [CrossRef] [Green Version]
- Padayatty, S.J.; Sun, H.; Wang, Y.; Riordan, H.D.; Hewitt, S.M.; Katz, A.; Wesley, R.A.; Levine, M. Vitamin C pharmacokinetics: Implications for oral and intravenous use. Ann. Intern. Med. 2004, 140, 533–537. [Google Scholar] [CrossRef]
- Wilson, J.X. Mechanism of action of vitamin C in sepsis: Ascorbate modulates redox signaling in endothelium. Biofactors 2009, 35, 5–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, J.X. Evaluation of vitamin C for adjuvant sepsis therapy. Antioxid. Redox Signal. 2013, 19, 2129–2140. [Google Scholar] [CrossRef]
- Frei, B.; Stocker, R.; England, L.; Ames, B.N. Ascorbate: The most effective antioxidant in human blood plasma. Adv. Exp. Med. Biol. 1990, 264, 155–163. [Google Scholar] [PubMed]
- Koekkoek, W.A.C.K.; van Zanten, A.R.H. Antioxidant vitamins and trace elements in critical illness. Nutr. Clin. Pract. 2016, 31, 457–474. [Google Scholar] [CrossRef] [PubMed]
- Nathens, A.B.; Neff, M.J.; Jurkovich, G.J.; Klotz, P.; Farver, K.; Ruzinski, J.T.; Radella, F.; Garcia, I.; Maier, R.V. Randomized, prospective trial of antioxidant supplementation in critically ill surgical patients. Ann. Surg. 2002, 236, 814–822. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.T.; Lin, H.I.; Yeh, D.Y.; Chou, T.Y.; Chen, C.F.; Leu, F.C.; Wang, D.; Hu, R.T. Effects of the antioxidants lycium barbarum and ascorbic acid on reperfusion liver injury in rats. Transpl. Proc. 2009, 41, 4110–4113. [Google Scholar] [CrossRef]
- Lee, W.-Y.; Lee, J.-S.; Lee, S.-M. Protective effects of combined ischemic preconditioning and ascorbic acid on mitochondrial injury in hepatic ischemia/reperfusion. J. Surg. Res. 2007, 142, 45–52. [Google Scholar] [CrossRef]
- Berger, M.M.; Soguel, L.; Shenkin, A.; Revelly, J.-P.; Pinget, C.; Baines, M.; Chioléro, R.L. Influence of early antioxidant supplements on clinical evolution and organ function in critically ill cardiac surgery, major trauma, and subarachnoid hemorrhage patients. Crit. Care 2008, 12, R101. [Google Scholar] [CrossRef] [Green Version]
- Oudemans-van Straaten, H.M.; Spoelstra-de Man, A.M.; de Waard, M.C. Vitamin C revisited. Crit. Care 2014, 18, 460. [Google Scholar] [CrossRef] [Green Version]
- Carr, A.C.; Shaw, G.M.; Fowler, A.A.; Natarajan, R. Ascorbate-dependent vasopressor synthesis: A rationale for vitamin C administration in severe sepsis and septic shock? Crit. Care 2015, 19, 418. [Google Scholar] [CrossRef] [Green Version]
- Kagan, V.E.; Serbinova, E.A.; Forte, T.; Scita, G.; Packer, L. Recycling of vitamin E in human low density lipoproteins. J. Lipid Res. 1992, 33, 385–397. [Google Scholar] [PubMed]
- Noguchi, N.; Iwaki, Y.; Takahashi, M.; Komuro, E.; Kato, Y.; Tamura, K.; Cynshi, O.; Kodama, T.; Niki, E. 2,3-Dihydro-5-hydroxy-2,2-dipentyl-4,6-di-tert-butylbenzofuran: Design and evaluation as a novel radical-scavenging antioxidant against lipid peroxidation. Arch. Biochem. Biophys. 1997, 342, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Eipper, B.A.; Mains, R.E. The role of ascorbate in the biosynthesis of neuroendocrine peptides. Am. J. Clin. Nutr. 1991, 54, 1153S–1156S. [Google Scholar] [CrossRef]
- Arrigoni, O.; De Tullio, M.C. The role of ascorbic acid in cell metabolism: Between gene-directed functions and unpredictable chemical reactions. J. Plant Physiol. 2000, 157, 481–488. [Google Scholar] [CrossRef]
- Gorres, K.L.; Raines, R.T. Prolyl 4-hydroxylase. Crit. Rev. Biochem. Mol. Biol. 2010, 45, 106–124. [Google Scholar] [CrossRef]
- Kivirikko, K.I.; Myllylä, R. Post-translational processing of procollagens. Ann. N. Y. Acad. Sci. 1985, 460, 187–201. [Google Scholar] [CrossRef]
- Kishimoto, Y.; Saito, N.; Kurita, K.; Shimokado, K.; Maruyama, N.; Ishigami, A. Ascorbic acid enhances the expression of type 1 and type 4 collagen and SVCT2 in cultured human skin fibroblasts. Biochem. Biophys. Res. Commun. 2013, 430, 579–584. [Google Scholar] [CrossRef]
- Nytko, K.J.; Maeda, N.; Schläfli, P.; Spielmann, P.; Wenger, R.H.; Stiehl, D.P. Vitamin C is dispensable for oxygen sensing in vivo. Blood 2011, 117, 5485–5493. [Google Scholar] [CrossRef] [Green Version]
- Bruick, R.K.; McKnight, S.L. A conserved family of prolyl-4-hydroxylases that modify HIF. Science 2001, 294, 1337–1340. [Google Scholar] [CrossRef] [Green Version]
- Grano, A.; De Tullio, M.C. Ascorbic acid as a sensor of oxidative stress and a regulator of gene expression: The Yin and Yang of vitamin C. Med. Hypotheses 2007, 69, 953–954. [Google Scholar] [CrossRef]
- Ke, Q.; Costa, M. Hypoxia-inducible factor-1 (HIF-1). Mol. Pharm. 2006, 70, 1469–1480. [Google Scholar] [CrossRef] [PubMed]
- Nelson, P.J.; Pruitt, R.E.; Henderson, L.L.; Jenness, R.; Henderson, L.M. Effect of ascorbic acid deficiency on the in vivo synthesis of carnitine. Biochim. Biophys. Acta (BBA)-Gen. Subj. 1981, 672, 123–127. [Google Scholar] [CrossRef]
- Rebouche, C.J. Ascorbic acid and carnitine biosynthesis. Am. J. Clin. Nutr. 1991, 54, 1147S–1152S. [Google Scholar] [CrossRef] [PubMed]
- Winter, B.K.; Fiskum, G.; Gallo, L.L. Effects of L-carnitine on serum triglyceride and cytokine levels in rat models of cachexia and septic shock. Br. J. Cancer 1995, 72, 1173–1179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foroozanfar, N.; Lucas, C.F.; Joss, D.V.; Hugh-Jones, K.; Hobbs, J.R. Ascorbate (1g/day) does not help the phagocyte killing defect of X-linked chronic granulomatous disease. Clin. Exp. Immunol. 1983, 51, 99–102. [Google Scholar]
- Goldschmidt, M.C. Reduced bactericidal activity in neutrophils from scorbutic animals and the effect of ascorbic acid on these target bacteria in vivo and in vitro. Am. J. Clin. Nutr. 1991, 54, 1214S–1220S. [Google Scholar] [CrossRef] [Green Version]
- Goldschmidt, M.C.; Masin, W.J.; Brown, L.R.; Wyde, P.R. The effect of ascorbic acid deficiency on leukocyte phagocytosis and killing of actinomyces viscosus. Int. J. Vitam. Nutr. Res. 1988, 58, 326–334. [Google Scholar]
- Rebora, A.; Dallegri, F.; Patrone, F. Neutrophil dysfunction and repeated infections: Influence of levamisole and ascorbic acid. Br. J. Dermatol. 1980, 102, 49–56. [Google Scholar] [CrossRef]
- Shilotri, P.G. Glycolytic, hexose monophosphate shunt and bactericidal activities of leukocytes in ascorbic acid deficient guinea pigs. J. Nutr. 1977, 107, 1507–1512. [Google Scholar] [CrossRef] [Green Version]
- Johnston, C.S.; Huang, S.N. Effect of ascorbic acid nutriture on blood histamine and neutrophil chemotaxis in guinea pigs. J. Nutr. 1991, 121, 126–130. [Google Scholar] [CrossRef] [Green Version]
- Carr, A.C.; Maggini, S. Vitamin C and immune function. Nutrients 2017, 9, 1211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Washko, P.W.; Wang, Y.; Levine, M. Ascorbic acid recycling in human neutrophils. J. Biol. Chem. 1993, 268, 15531–15535. [Google Scholar] [PubMed]
- Chen, Y.; Luo, G.; Yuan, J.; Wang, Y.; Yang, X.; Wang, X.; Li, G.; Liu, Z.; Zhong, N. Vitamin C mitigates oxidative stress and tumor necrosis factor-alpha in severe community-acquired pneumonia and LPS-induced macrophages. Mediat. Inflamm. 2014, 2014, 426740. [Google Scholar] [CrossRef] [Green Version]
- Cárcamo, J.M.; Pedraza, A.; Bórquez-Ojeda, O.; Zhang, B.; Sanchez, R.; Golde, D.W. Vitamin C is a kinase inhibitor: Dehydroascorbic acid inhibits IkappaBalpha kinase beta. Mol. Cell. Biol. 2004, 24, 6645–6652. [Google Scholar] [CrossRef] [Green Version]
- Peng, Y.; Kwok, K.H.H.; Yang, P.-H.; Ng, S.S.M.; Liu, J.; Wong, O.G.; He, M.-L.; Kung, H.-F.; Lin, M.C.M. Ascorbic acid inhibits ROS production, NF-kappa B activation and prevents ethanol-induced growth retardation and microencephaly. Neuropharmacology 2005, 48, 426–434. [Google Scholar] [CrossRef]
- Ferrón-Celma, I.; Mansilla, A.; Hassan, L.; Garcia-Navarro, A.; Comino, A.-M.; Bueno, P.; Ferrón, J.-A. Effect of vitamin C administration on neutrophil apoptosis in septic patients after abdominal surgery. J. Surg. Res. 2009, 153, 224–230. [Google Scholar] [CrossRef]
- Hampton, M.B.; Fadeel, B.; Orrenius, S. Redox regulation of the caspases during apoptosis. Ann. N. Y. Acad. Sci. 1998, 854, 328–335. [Google Scholar] [CrossRef]
- Mikacenic, C.; Moore, R.; Dmyterko, V.; West, T.E.; Altemeier, W.A.; Liles, W.C.; Lood, C. Neutrophil extracellular traps (NETs) are increased in the alveolar spaces of patients with ventilator-associated pneumonia. Crit. Care 2018, 22, 358. [Google Scholar] [CrossRef] [Green Version]
- Zawrotniak, M.; Rapala-Kozik, M. Neutrophil extracellular traps (NETs)-formation and implications. Acta Biochim. Pol. 2013, 60, 277–284. [Google Scholar] [CrossRef] [Green Version]
- Lefrançais, E.; Mallavia, B.; Zhuo, H.; Calfee, C.S.; Looney, M.R. Maladaptive role of neutrophil extracellular traps in pathogen-induced lung injury. JCI Insight 2018, 3, 3. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Qi, H.; Kan, K.; Chen, J.; Xie, H.; Guo, X.; Zhang, L. Neutrophil extracellular traps promote hypercoagulability in patients with sepsis. Shock 2017, 47, 132–139. [Google Scholar] [CrossRef]
- Liu, S.; Su, X.; Pan, P.; Zhang, L.; Hu, Y.; Tan, H.; Wu, D.; Liu, B.; Li, H.; Li, H.; et al. Neutrophil extracellular traps are indirectly triggered by lipopolysaccharide and contribute to acute lung injury. Sci. Rep. 2016, 6, 37252. [Google Scholar] [CrossRef] [Green Version]
- Czaikoski, P.G.; Mota, J.M.S.C.; Nascimento, D.C.; Sônego, F.; Castanheira, F.V.E.S.; Melo, P.H.; Scortegagna, G.T.; Silva, R.L.; Barroso-Sousa, R.; Souto, F.O.; et al. Neutrophil Extracellular Traps Induce Organ Damage during Experimental and Clinical Sepsis. PLoS ONE 2016, 11, e0148142. [Google Scholar] [CrossRef] [Green Version]
- Saffarzadeh, M.; Juenemann, C.; Queisser, M.A.; Lochnit, G.; Barreto, G.; Galuska, S.P.; Lohmeyer, J.; Preissner, K.T. Neutrophil extracellular traps directly induce epithelial and endothelial cell death: A predominant role of histones. PLoS ONE 2012, 7, e32366. [Google Scholar] [CrossRef]
- Hirose, T.; Hamaguchi, S.; Matsumoto, N.; Irisawa, T.; Seki, M.; Tasaki, O.; Hosotsubo, H.; Yamamoto, N.; Yamamoto, K.; Akeda, Y.; et al. Presence of neutrophil extracellular traps and citrullinated histone H3 in the bloodstream of critically ill patients. PLoS ONE 2014, 9, e111755. [Google Scholar] [CrossRef] [Green Version]
- Kaplan, M.J.; Radic, M. Neutrophil extracellular traps: Double-edged swords of innate immunity. J. Immunol. 2012, 189, 2689–2695. [Google Scholar] [CrossRef] [Green Version]
- Sørensen, O.E.; Borregaard, N. Neutrophil extracellular traps - the dark side of neutrophils. J. Clin. Investig. 2016, 126, 1612–1620. [Google Scholar] [CrossRef] [Green Version]
- Maruchi, Y.; Tsuda, M.; Mori, H.; Takenaka, N.; Gocho, T.; Huq, M.A.; Takeyama, N. Plasma myeloperoxidase-conjugated DNA level predicts outcomes and organ dysfunction in patients with septic shock. Crit. Care 2018, 22, 176. [Google Scholar] [CrossRef] [Green Version]
- Manda, A.; Pruchniak, M.P.; Araźna, M.; Demkow, U.A. Neutrophil extracellular traps in physiology and pathology. Cent. Eur. J. Immunol. 2014, 39, 116–121. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, X.M.; Biron, B.M.; Reichner, J.S. Consequences of extracellular trap formation in sepsis. Curr. Opin. Hematol. 2017, 24, 66–71. [Google Scholar] [CrossRef] [Green Version]
- Mohammed, B.M.; Fisher, B.J.; Kraskauskas, D.; Farkas, D.; Brophy, D.F.; Fowler, A.A.; Natarajan, R. Vitamin C: A novel regulator of neutrophil extracellular trap formation. Nutrients 2013, 5, 3131–3151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamaguchi, S.; Akeda, Y.; Yamamoto, N.; Seki, M.; Yamamoto, K.; Oishi, K.; Tomono, K. Origin of circulating free DNA in sepsis: Analysis of the CLP mouse model. Mediat. Inflamm. 2015, 2015, 614518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakahira, K.; Kyung, S.-Y.; Rogers, A.J.; Gazourian, L.; Youn, S.; Massaro, A.F.; Quintana, C.; Osorio, J.C.; Wang, Z.; Zhao, Y.; et al. Circulating mitochondrial DNA in patients in the ICU as a marker of mortality: Derivation and validation. PLoS Med. 2013, 10, e1001577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manning, J.; Mitchell, B.; Appadurai, D.A.; Shakya, A.; Pierce, L.J.; Wang, H.; Nganga, V.; Swanson, P.C.; May, J.M.; Tantin, D.; et al. Vitamin C promotes maturation of T-cells. Antioxid. Redox Signal. 2013, 19, 2054–2067. [Google Scholar] [CrossRef] [Green Version]
- Huijskens, M.J.A.J.; Walczak, M.; Koller, N.; Briedé, J.J.; Senden-Gijsbers, B.L.M.G.; Schnijderberg, M.C.; Bos, G.M.J.; Germeraad, W.T.V. Technical advance: Ascorbic acid induces development of double-positive T cells from human hematopoietic stem cells in the absence of stromal cells. J. Leukoc. Biol. 2014, 96, 1165–1175. [Google Scholar] [CrossRef]
- Agathocleous, M.; Meacham, C.E.; Burgess, R.J.; Piskounova, E.; Zhao, Z.; Crane, G.M.; Cowin, B.L.; Bruner, E.; Murphy, M.M.; Chen, W.; et al. Ascorbate regulates haematopoietic stem cell function and leukaemogenesis. Nature 2017, 549, 476–481. [Google Scholar] [CrossRef]
- Lee Chong, T.; Ahearn, E.L.; Cimmino, L. Reprogramming the epigenome with vitamin C. Front. Cell Dev. Biol. 2019, 7, 128. [Google Scholar] [CrossRef]
- Young, J.I.; Züchner, S.; Wang, G. Regulation of the epigenome by vitamin C. Annu. Rev. Nutr. 2015, 35, 545–564. [Google Scholar] [CrossRef] [Green Version]
- Kallio, J.; Jaakkola, M.; Mäki, M.; Kilpeläinen, P.; Virtanen, V. Vitamin C inhibits staphylococcus aureus growth and enhances the inhibitory effect of quercetin on growth of Escherichia coli in vitro. Planta Med. 2012, 78, 1824–1830. [Google Scholar] [CrossRef] [Green Version]
- Rawal, B.D. Bactericidal action of ascorbic acid on Psuedomonas aeruginosa: Alteration of cell surface as a possible mechanism. Chemotherapy 1978, 24, 166–171. [Google Scholar] [CrossRef]
- Mikirova, N.; Riordan, N.; Casciari, J. Modulation of cytokines in cancer patients by intravenous ascorbate therapy. Med. Sci. Monit. 2016, 22, 14–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borrelli, E.; Roux-Lombard, P.; Grau, G.E.; Girardin, E.; Ricou, B.; Dayer, J.; Suter, P.M. Plasma concentrations of cytokines, their soluble receptors, and antioxidant vitamins can predict the development of multiple organ failure in patients at risk. Crit. Care Med. 1996, 24, 392–397. [Google Scholar] [CrossRef] [PubMed]
- Hamacher, J.; Hadizamani, Y.; Borgmann, M.; Mohaupt, M.; Männel, D.N.; Moehrlen, U.; Lucas, R.; Stammberger, U. Cytokine-Ion Channel Interactions in Pulmonary Inflammation. Front. Immunol. 2017, 8, 1644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.; Fu, G.; Yao, Q.; Cheng, G. Relation of thrombomodulin, TFPI and plasma antioxidants in healthy individuals and patients with coronary heart disease. Acta Cardiol. 2008, 63, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Puskarich, M.A.; Cornelius, D.C.; Tharp, J.; Nandi, U.; Jones, A.E. Plasma syndecan-1 levels identify a cohort of patients with severe sepsis at high risk for intubation after large-volume intravenous fluid resuscitation. J. Crit. Care 2016, 36, 125–129. [Google Scholar] [CrossRef]
- Hayashida, K.; Parks, W.C.; Park, P.W. Syndecan-1 shedding facilitates the resolution of neutrophilic inflammation by removing sequestered CXC chemokines. Blood 2009, 114, 3033–3043. [Google Scholar] [CrossRef] [Green Version]
- Smart, L.; Bosio, E.; Macdonald, S.P.J.; Dull, R.; Fatovich, D.M.; Neil, C.; Arendts, G. Glycocalyx biomarker syndecan-1 is a stronger predictor of respiratory failure in patients with sepsis due to pneumonia, compared to endocan. J. Crit. Care 2018, 47, 93–98. [Google Scholar] [CrossRef]
- Wu, F.; Peng, Z.; Park, P.W.; Kozar, R.A. Loss of Syndecan-1 Abrogates the Pulmonary Protective Phenotype Induced by Plasma After Hemorrhagic Shock. Shock 2017, 48, 340–345. [Google Scholar] [CrossRef]
- Palaiologou, M.; Delladetsima, I.; Tiniakos, D. CD138 (syndecan-1) expression in health and disease. Histol. Histopathol. 2014, 29, 177–189. [Google Scholar]
- Sallisalmi, M.; Tenhunen, J.; Yang, R.; Oksala, N.; Pettilä, V. Vascular adhesion protein-1 and syndecan-1 in septic shock. Acta Anaesthesiol. Scand. 2012, 56, 316–322. [Google Scholar] [CrossRef]
- Wei, S.; Gonzalez Rodriguez, E.; Chang, R.; Holcomb, J.B.; Kao, L.S.; Wade, C.E. PROPPR Study Group Elevated Syndecan-1 after Trauma and Risk of Sepsis: A Secondary Analysis of Patients from the Pragmatic, Randomized Optimal Platelet and Plasma Ratios (PROPPR) Trial. J. Am. Coll. Surg. 2018, 227, 587–595. [Google Scholar] [CrossRef] [PubMed]
- Nelson, A.; Johansson, J.; Tydén, J.; Bodelsson, M. Circulating syndecans during critical illness. APMIS 2017, 125, 468–475. [Google Scholar] [CrossRef] [PubMed]
- Bertrand, J.; Bollmann, M. Soluble syndecans: Biomarkers for diseases and therapeutic options. Br. J. Pharmacol. 2019, 176, 67–81. [Google Scholar] [CrossRef] [PubMed]
- Murphy, L.S.; Wickersham, N.; McNeil, J.B.; Shaver, C.M.; May, A.K.; Bastarache, J.A.; Ware, L.B. Endothelial glycocalyx degradation is more severe in patients with non-pulmonary sepsis compared to pulmonary sepsis and associates with risk of ARDS and other organ dysfunction. Ann. Intensive Care 2017, 7, 102. [Google Scholar] [CrossRef]
- Nelson, A.; Berkestedt, I.; Schmidtchen, A.; Ljunggren, L.; Bodelsson, M. Increased levels of glycosaminoglycans during septic shock: Relation to mortality and the antibacterial actions of plasma. Shock 2008, 30, 623–627. [Google Scholar] [CrossRef]
- Kashiouris, M.G.; Qiao, X.; Fisher, B.J.; Truwit, J.D.; Hite, R.D.; Morris, P.E.; Martin, G.S.; Fowler, A.A. A Multicenter Parsimonious Biomarker Mortality Prediction Model for Sepsis-Induced ARDS. In A95. Critical Care: Alice in Wonderland—Biomarkers in Critical Illness: Clues to be Unravelled; American Thoracic Society: New York, NY, USA, 2019; p. A2381. [Google Scholar]
- Qiao, X.; Fisher, B.; Kashiouris, M.G.; Truwit, J.D.; Hite, R.D.; Morris, P.E.; Martin, G.S.; Fowler, A.A. Effects of High Dose Intravenous Vitamin C (IVC) on Plasma Cell-Free DNA Levels in Patients with Sepsis-Associated ARDS. In A60. Lung Injury, Sepsis, and Ards; American Thoracic Society: New York, NY, USA, 2019; p. A2100. [Google Scholar]
- Pignatelli, P.; Sanguigni, V.; Paola, S.G.; Lo Coco, E.; Lenti, L.; Violi, F. Vitamin C inhibits platelet expression of CD40 ligand. Free Radic. Biol. Med. 2005, 38, 1662–1666. [Google Scholar] [CrossRef] [Green Version]
- Horrobin, D.F. Ascorbic acid and prostaglandin synthesis. Subcell. Biochem. 1996, 25, 109–115. [Google Scholar]
- Mohammed, B.M.; Sanford, K.W.; Fisher, B.J.; Martin, E.J.; Contaifer, D.; Warncke, U.O.; Wijesinghe, D.S.; Chalfant, C.E.; Brophy, D.F.; Fowler Iii, A.A.; et al. Impact of high dose vitamin C on platelet function. World J. Crit. Care Med. 2017, 6, 37–47. [Google Scholar] [CrossRef]
- Cudjoe, E.K.; Hassan, Z.H.; Kang, L.; Reynolds, P.S.; Fisher, B.J.; McCarter, J.; Sweeney, C.; Martin, E.J.; Middleton, P.; Ellenberg, M.; et al. Temporal map of the pig polytrauma plasma proteome with fluid resuscitation and intravenous vitamin C treatment. J. Thromb. Haemost. 2019, 17, 1827–1837. [Google Scholar] [CrossRef]
- Galley, H.F.; Davies, M.J.; Webster, N.R. Ascorbyl radical formation in patients with sepsis: Effect of ascorbate loading. Free Radic. Biol. Med. 1996, 20, 139–143. [Google Scholar] [CrossRef]
- Fowler, A.A.; Syed, A.A.; Knowlson, S.; Sculthorpe, R.; Farthing, D.; De Wilde, C.; Farthing, C.A.; Larus, T.L.; Martin, E.; Brophy, D.F.; et al. Medical Respiratory Intensive Care Unit Nursing, Fisher BJ, Natarajan R. Phase I safety trial of intravenous ascorbic acid in patients with severe sepsis. J. Transl. Med. 2014, 12, 32. [Google Scholar] [CrossRef] [Green Version]
- Seno, T.; Inoue, N.; Matsui, K.; Ejiri, J.; Hirata, K.-I.; Kawashima, S.; Yokoyama, M. Functional expression of sodium-dependent vitamin C transporter 2 in human endothelial cells. J. Vasc. Res. 2004, 41, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Oudemans-van Straaten, H.M.; Elbers, P.W.G.; Spoelstra-de Man, A.M.E. How to give vitamin C a cautious but fair chance in severe sepsis. Chest 2017, 151, 1199–1200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zabet, M.H.; Mohammadi, M.; Ramezani, M.; Khalili, H. Effect of high-dose Ascorbic acid on vasopressor’s requirement in septic shock. J. Res. Pharm. Pract. 2016, 5, 94–100. [Google Scholar] [PubMed]
- Bürzle, M.; Hediger, M.A. Functional and physiological role of vitamin C transporters. Curr. Top. Membr. 2012, 70, 357–375. [Google Scholar]
- Abrams, S.T.; Morton, B.; Alhamdi, Y.; Alsabani, M.; Lane, S.; Welters, I.D.; Wang, G.; Toh, C.-H. A novel assay for neutrophil extracellular trap formation independently predicts disseminated intravascular coagulation and mortality in critically ill patients. Am. J. Respir. Crit. Care Med. 2019, 200, 869–880. [Google Scholar] [CrossRef]
- Gupta, A.K.; Joshi, M.B.; Philippova, M.; Erne, P.; Hasler, P.; Hahn, S.; Resink, T.J. Activated endothelial cells induce neutrophil extracellular traps and are susceptible to NETosis-mediated cell death. FEBS Lett. 2010, 584, 3193–3197. [Google Scholar] [CrossRef] [Green Version]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil extracellular traps kill bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef]
- Bozonet, S.M.; Carr, A.C. The Role of Physiological Vitamin C Concentrations on Key Functions of Neutrophils Isolated from Healthy Individuals. Nutrients 2019, 11, 1363. [Google Scholar] [CrossRef] [Green Version]
- May, J.M.; Harrison, F.E. Role of vitamin C in the function of the vascular endothelium. Antioxid. Redox Signal. 2013, 19, 2068–2083. [Google Scholar] [CrossRef] [Green Version]
- Fisher, B.J.; Seropian, I.M.; Kraskauskas, D.; Thakkar, J.N.; Voelkel, N.F.; Fowler, A.A.; Natarajan, R. Ascorbic acid attenuates lipopolysaccharide-induced acute lung injury. Crit. Care Med. 2011, 39, 1454–1460. [Google Scholar] [CrossRef] [PubMed]
- Burchfield, J.S.; Li, Q.; Wang, H.Y.; Wang, R.-F. JMJD3 as an epigenetic regulator in development and disease. Int. J. Biochem. Cell Biol. 2015, 67, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Brogdon, J.L.; Xu, Y.; Szabo, S.J.; An, S.; Buxton, F.; Cohen, D.; Huang, Q. Histone deacetylase activities are required for innate immune cell control of Th1 but not Th2 effector cell function. Blood 2007, 109, 1123–1130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, L.D.; Le, T.; Fan, G. DNA Methylation and Its Basic Function. Neuropsychopharmacology 2013, 38, 23–38. [Google Scholar] [CrossRef] [Green Version]
- Intravenous Vitamin C Employed as Adjunctive Therapy in Recurrent Acute Respiratory Distress Syndrome (ARDS): A Case Report A54. Critical Care Case Reports: Acute Hypoxemic Respiratory Failure/Ards. Available online: https://www.atsjournals.org/doi/abs/10.1164/ajrccm-conference.2016.193.1_MeetingAbstracts.A1892 (accessed on 24 September 2019).
- Fisher, B.J.; Kraskauskas, D.; Martin, E.J.; Farkas, D.; Wegelin, J.A.; Brophy, D.; Ward, K.R.; Voelkel, N.F.; Fowler, A.A.; Natarajan, R. Mechanisms of attenuation of abdominal sepsis induced acute lung injury by ascorbic acid. Am. J. Physiol. Lung Cell Mol. Physiol. 2012, 303, L20–L32. [Google Scholar] [CrossRef]
- Sawyer, M.A.J.; Mike, J.J.; Chavin, K.; Marino, P.L. Antioxidant therapy and survival in ARDS. Crit. Care Med. 1989, 17, S153. [Google Scholar]
- Vincent, J.L.; Moreno, R.; Takala, J.; Willatts, S.; De Mendonça, A.; Bruining, H.; Reinhart, C.K.; Suter, P.; Thijs, L.G. The SOFA (Sepsis-related Organ Failure Assessment) score to describe organ dysfunction/failure. Intensive Care Med. 1996, 22, 707–710. [Google Scholar] [CrossRef]
- Marik, P.E.; Khangoora, V.; Rivera, R.; Hooper, M.H.; Catravas, J. Hydrocortisone, Vitamin C, and Thiamine for the Treatment of Severe Sepsis and Septic Shock: A Retrospective Before-After Study. Chest 2017, 151, 1229–1238. [Google Scholar] [CrossRef]
- Li, J. Evidence is stronger than you think: A meta-analysis of vitamin C use in patients with sepsis. Crit. Care 2018, 22, 258. [Google Scholar] [CrossRef] [Green Version]
- Fowler, A.A.; Truwit, J.D.; Hite, R.D.; Morris, P.E.; DeWilde, C.; Priday, A.; Fisher, B.; Thacker, L.R.; Natarajan, R.; Brophy, D.F.; et al. Effect of Vitamin C Infusion on Organ Failure and Biomarkers of Inflammation and Vascular Injury in Patients With Sepsis and Severe Acute Respiratory Failure: The CITRIS-ALI Randomized Clinical Trial. JAMA 2019, 322, 1261–1270. [Google Scholar] [CrossRef]
- Tanaka, H.; Matsuda, T.; Miyagantani, Y.; Yukioka, T.; Matsuda, H.; Shimazaki, S. Reduction of resuscitation fluid volumes in severely burned patients using ascorbic acid administration: A randomized, prospective study. Arch. Surg. 2000, 135, 326–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hager, D.N.; Martin, G.S.; Sevransky, J.E.; Hooper, M.H. Glucometry when using vitamin C in sepsis: A note of caution. Chest 2018, 154, 228–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sartor, Z.; Kesey, J.; Dissanaike, S. The effects of intravenous vitamin C on point-of-care glucose monitoring. J. Burn Care Res. 2015, 36, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.E.; Brown, C.S.; Manning, B.M.; May, T.; Riker, R.R.; Lerwick, P.A.; Hayes, T.L.; Fraser, G.L. Accuracy of Point-of-Care Blood Glucose Level Measurements in Critically Ill Patients with Sepsis Receiving High-Dose Intravenous Vitamin, C. Pharmacotherapy 2018, 38, 1155–1161. [Google Scholar] [CrossRef]
Figure 1.
The last step in vitamin C (C6H8O6, or l-Ascorbic acid) biosynthesis. Humans have lost the ability to synthesize the GLO enzyme, and thus are dependent on exogenous vitamin C intake through their diet. Modified from: U.S. National Library of Medicine, PubChem.
Figure 1.
The last step in vitamin C (C6H8O6, or l-Ascorbic acid) biosynthesis. Humans have lost the ability to synthesize the GLO enzyme, and thus are dependent on exogenous vitamin C intake through their diet. Modified from: U.S. National Library of Medicine, PubChem.

Figure 2.
Ascorbic Acid (AA) and dehydroascorbic Acid (DHA) transporters (left). Concentration of AA and DHA in human organs and cells in uM (right). Inspired by Padayatty and Levine [25].
Figure 2.
Ascorbic Acid (AA) and dehydroascorbic Acid (DHA) transporters (left). Concentration of AA and DHA in human organs and cells in uM (right). Inspired by Padayatty and Levine [25].

Figure 3.
Infographic of differential vitamin C peak plasma concentrations based on alternative routes of administration and dosage.
Figure 3.
Infographic of differential vitamin C peak plasma concentrations based on alternative routes of administration and dosage.

Figure 4.
Pleiotropic effects of high-dose intravenous vitamin C (HDIVC) in sepsis-induced acute respiratory distress syndrome (ARDS). The orange star [✯] points to possible therapeutic targets of HDIVC. The figure illustrates a human alveolus with the capillary membrane, and the blood-gas barrier during sepsis.
Figure 4.
Pleiotropic effects of high-dose intravenous vitamin C (HDIVC) in sepsis-induced acute respiratory distress syndrome (ARDS). The orange star [✯] points to possible therapeutic targets of HDIVC. The figure illustrates a human alveolus with the capillary membrane, and the blood-gas barrier during sepsis.

Figure 5.
Kaplan–Meier mortality curves in patients with sepsis induced acute respiratory distress syndrome (ARDS) who were randomized to receive a 4-day course of high-dose intravenous vitamin C (HDIVC) versus placebo, upon ARDS onset-recognition.
Figure 5.
Kaplan–Meier mortality curves in patients with sepsis induced acute respiratory distress syndrome (ARDS) who were randomized to receive a 4-day course of high-dose intravenous vitamin C (HDIVC) versus placebo, upon ARDS onset-recognition.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Pleiotropic physiologic functions of vitamin C.
| The key antioxidant of the body | Reverses the oxidation of lipids by the neutrophil reactive oxygen species (ROS) [32,33]. Reduces depletion of other antioxidants (e.g., vitamin E and glutathione) to prevent oxidation of lipids, proteins and DNA [34,35,36,37,38,39,40]. |
| Norepinephrine biosynthesis | Cofactor for Dopamine ß-Hydroxylase, catalyzing the formation of norepinephrine from dopamine. Enhances adrenergic receptor activity [41]. |
| Dopamine biosynthesis | Facilitates recycling of the enzyme cofactor tetrahydrobiopterin (BH4); a required and rate-limiting step in the hydroxylation of l-tyrosine to form l-DOPA [42,43]. |
| Vasopressin biosynthesis | Cofactor for peptidylglycine α-amidating monooxygenase (PAM), involved in vasopressin biosynthesis [44]. |
| Connective tissue maintenance | Vital in wound healing; cofactor for Propyl 3-hydroxylase, prolyl 4-hydroxylase, and lysyl hydroxylase which catalyze the formation of procollagen and elastin biosynthesis [45,46]. Catalyzes the hydroxylation of procollagen to form the collagen triple-helix [47]. Induces fibroblast collagen gene expression, stimulating the production of new collagen [48]. |
| Regulation of cellular gene expression in response to hypoxia and stress | Needed for the hydroxylation (thus downregulation) of Hypoxia Induced Factor 1α (HIF-1α) by propyl and lysyl hydroxylases and FIF-1 (asparaginyl hydroxylase or factor inhibiting HIF-1) [49,50,51]. HIF-1α is a protein-transcription factor that regulates hundreds of genes in response to hypoxia and cellular stress, and is a marker of cellular hypoxia with increased expression in states of shock [52]. |
| Carnitine biosynthesis | Cofactor for γ-butyrobetaine hydroxylase, a dioxygenase involved in carnitine synthesis, which transports fatty acids into the mitochondria [53,54]. L-Carnitine can down-modulate tumor necrosis factor (TNF-α) by endotoxins, affect lipid metabolism, and reduce septic shock severity [55]. |
| Phagocytic cell function | Severe vitamin C deficiency (scorbutic) results in impaired neutrophilic phagocytosis and ROS generation [56,57,58,59,60]. In situations of impaired neutrophilic ROS production, vitamin C enhances the hexose monophosphate shunt (HMPS) and antibody dependent cell cytotoxicity (ADCC) resulting in increased bacterial killing [60]. Improves chemotaxis [61]. Accumulation in neutrophils may protect them from neutrophil dependent oxidative bursts [62,63]. Reduces inflammation and ROS via attenuation of NF-κB activation [64,65,66]. |
| Inflammation: Immune cell clearance | Promotes neutrophil apoptosis, instead of necrosis via activation of caspase-3 proteins [67,68]. High-dose intravenous vitamin C (HDIVC) treatment has been shown to decrease circulating plasma cell-free DNA (resulting from neutrophil extracellular trap (NET) formations, or NETosis), and have been implicated in sepsis-induced end-organ failure [69,70,71,72,73,74,75,76,77,78,79,80,81,82,83,84]. |
| Lymphocytic function | May promote lymphocytic proliferation, differentiation, and maturation [85,86]. |
| Epigenetic modulation | Cofactor for ten-eleven translocation (TET) enzymes and Jumonji-C domain-containing histone demethylases (JHDMs); vitamin C increasing enzymatic activity of both, resulting in increased DNA demethylation and histone demethylation, respectively, which controls gene transcription and gene activation or repression [87,88,89]. |
| Direct antimicrobial activity | High concentrations directly inhibit bacterial growth and exhibits bactericidal activity in vitro [90,91]. |
| Inflammatory mediators | Modulates cytokine production and can decrease circulating histamine levels [61,92,93,94]. |
| Endothelial function | HDIVC decreases circulating thrombomodulin, an endothelial membrane protein receptor for thrombin that converts thrombin to an anticoagulant capable of activating protein C [95]. Decreases plasma Syndecan-1 levels, a by-product of endothelial glycocalyx shedding [96,97,98,99,100,101,102,103,104,105,106,107,108]. |
| Platelet function and Thrombosis | Alters platelet oxidative states by inhibiting CD40 ligand expression on platelet surfaces [109]. Prolonged platelet exposure to HDIVC increases Thromboxane-B2 and Prostagladin-E2 levels [110,111]. HDIVC stabilizes ADAMTS13 levels and its von-Willebrand factor cleavage activity [112]. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kashiouris, M.G.; L’Heureux, M.; Cable, C.A.; Fisher, B.J.; Leichtle, S.W.; Fowler, A.A. The Emerging Role of Vitamin C as a Treatment for Sepsis. Nutrients 2020, 12, 292. https://doi.org/10.3390/nu12020292
AMA Style
Kashiouris MG, L’Heureux M, Cable CA, Fisher BJ, Leichtle SW, Fowler AA. The Emerging Role of Vitamin C as a Treatment for Sepsis. Nutrients. 2020; 12(2):292. https://doi.org/10.3390/nu12020292
Chicago/Turabian StyleKashiouris, Markos G., Michael L’Heureux, Casey A. Cable, Bernard J. Fisher, Stefan W. Leichtle, and Alpha A. Fowler. 2020. "The Emerging Role of Vitamin C as a Treatment for Sepsis" Nutrients 12, no. 2: 292. https://doi.org/10.3390/nu12020292
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.