
Severity of COVID-19 Patients Predicted by Serum Sphingolipids Signature

 , , , , , and
, , , , , and
Abstract
:1. Introduction
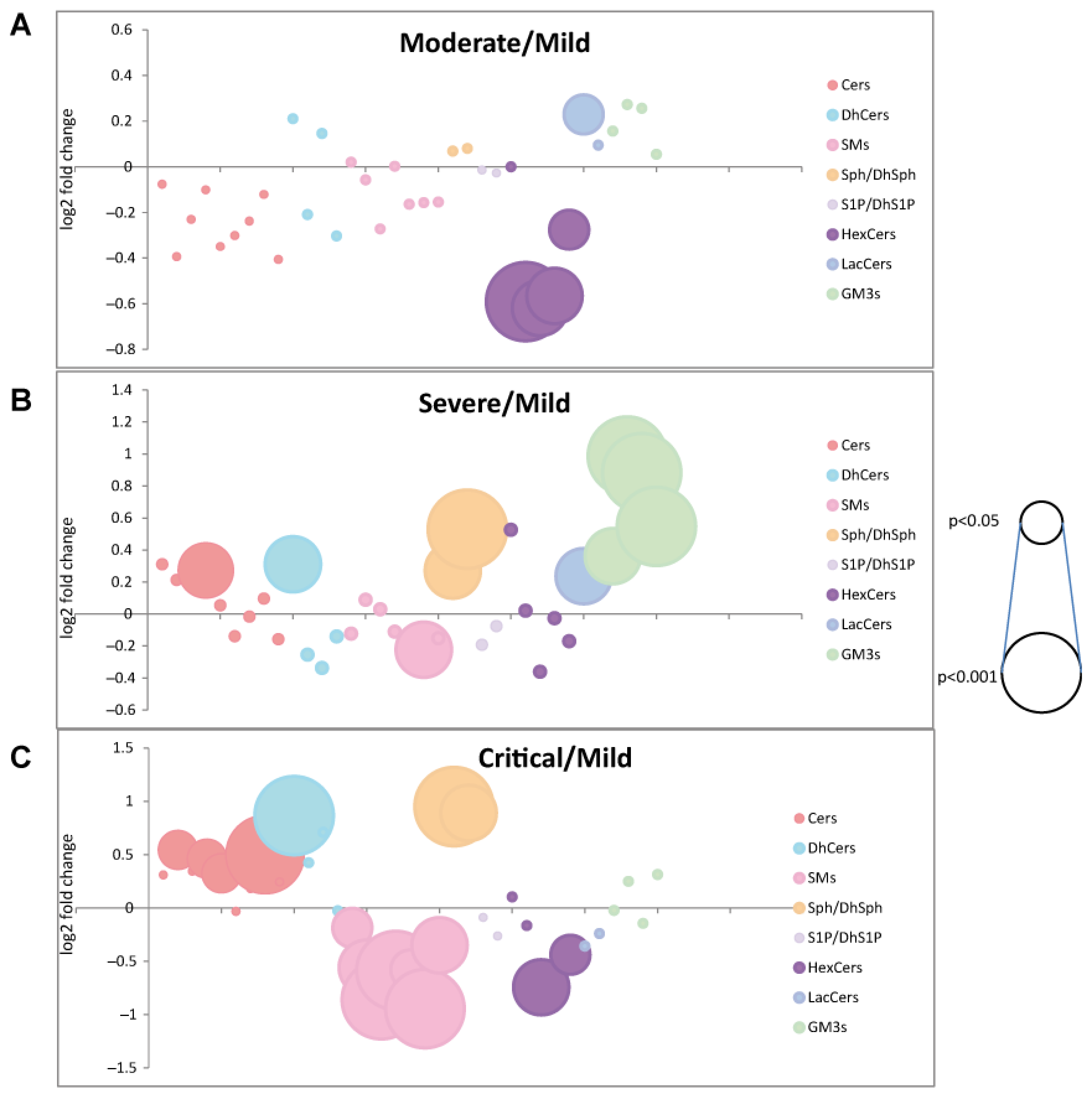
2. Results
2.1. Biochemical Parameters Assessment
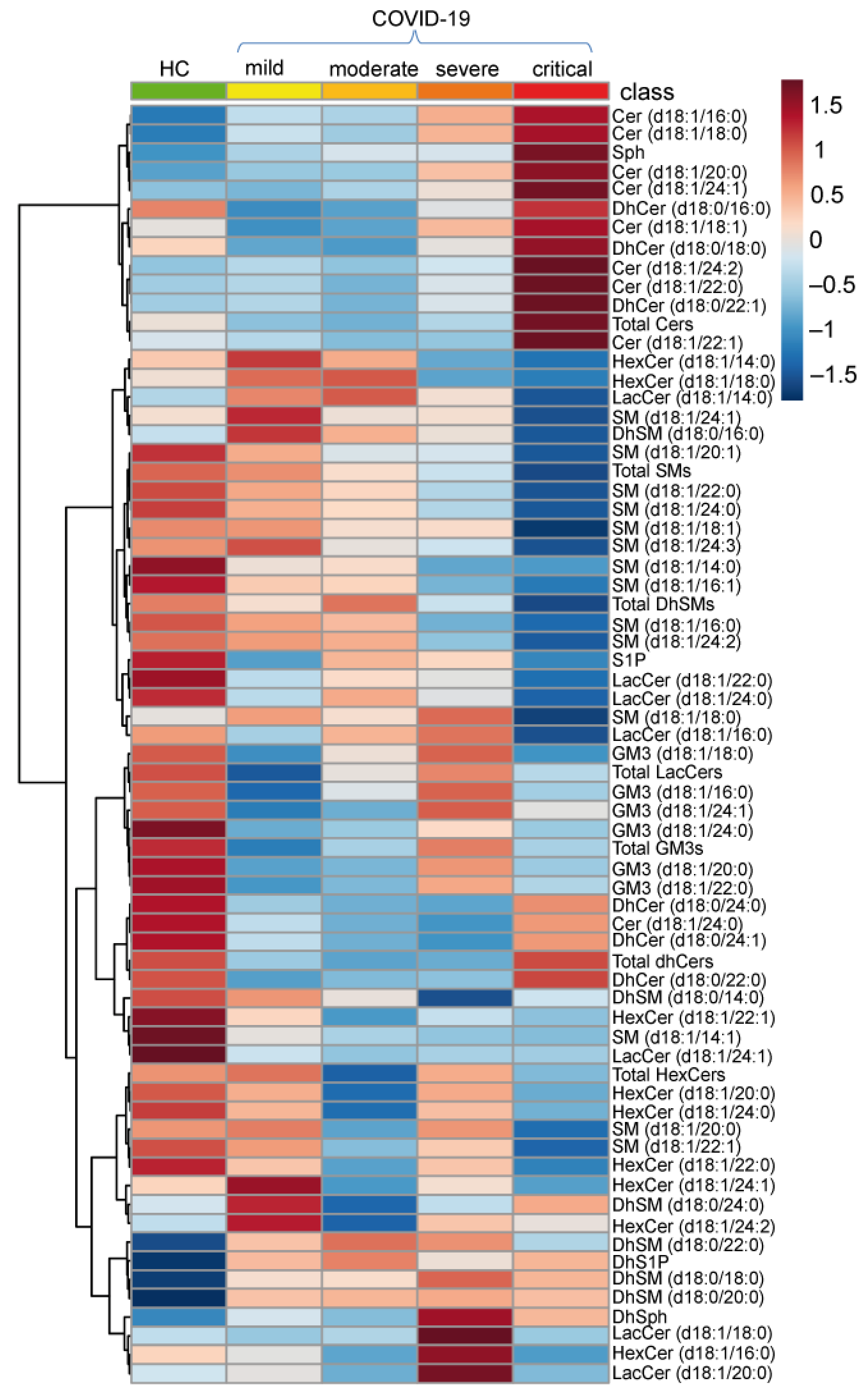
2.2. Serum Sphingolipid Levels Changes in COVID-19
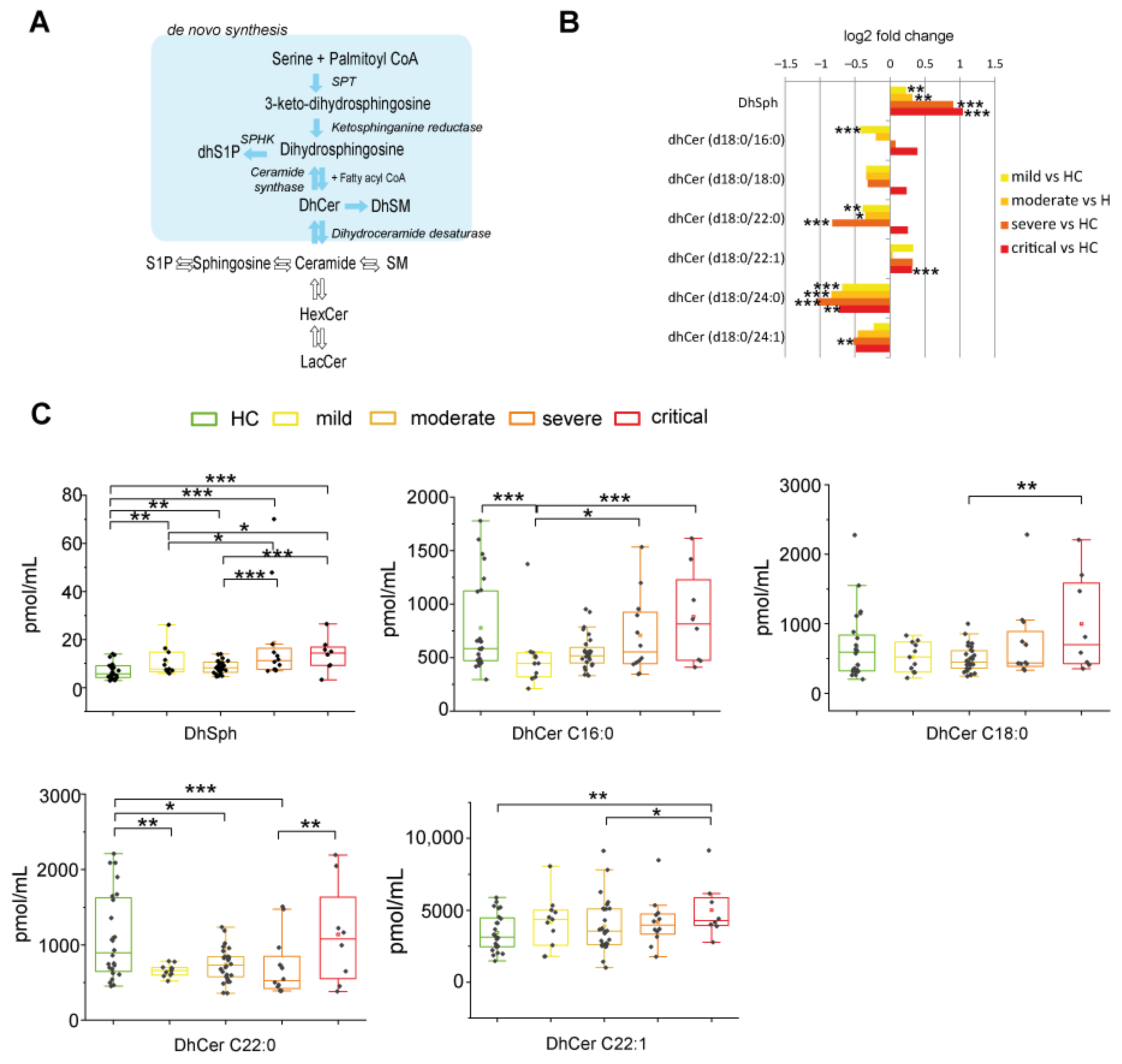
2.2.1. De Novo Pathway Ceramide Precursor, Dihydrosphingosine (DhSph), Increased with Disease Severity Whereas Dihydroceramides (DhCer) Levels Were at Variance According with COVID-19 Patient Severity and Acyl-Chain Length
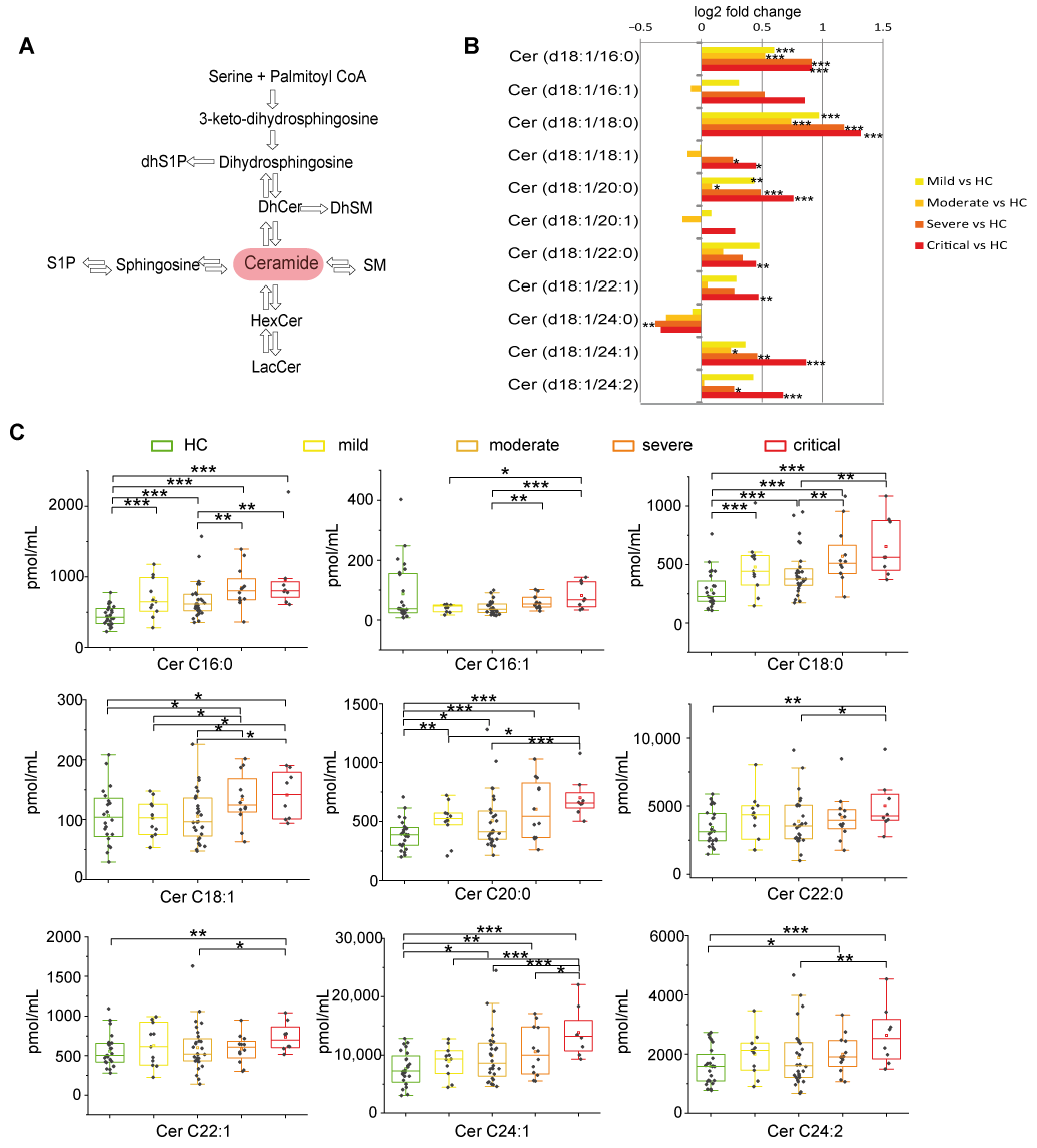
2.2.2. Long Chain and Very Long Chain Ceramides Progressively Increased with Disease Severity except for Ceramide C24:0
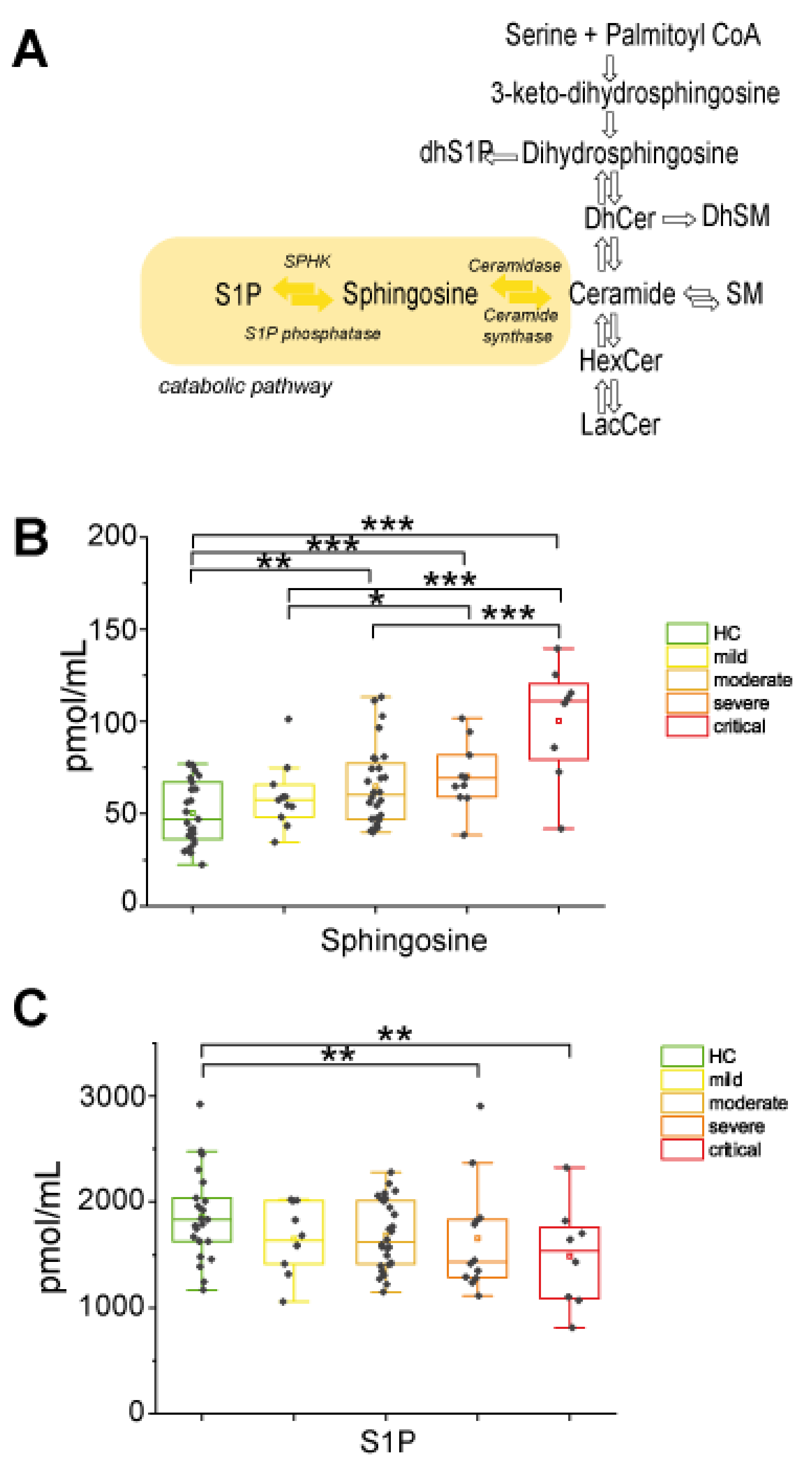
2.2.3. In the Catabolic Pathway, Sphingosine Increased with Disease Severity Whereas S1P Decreased in Severe and Critical Patients
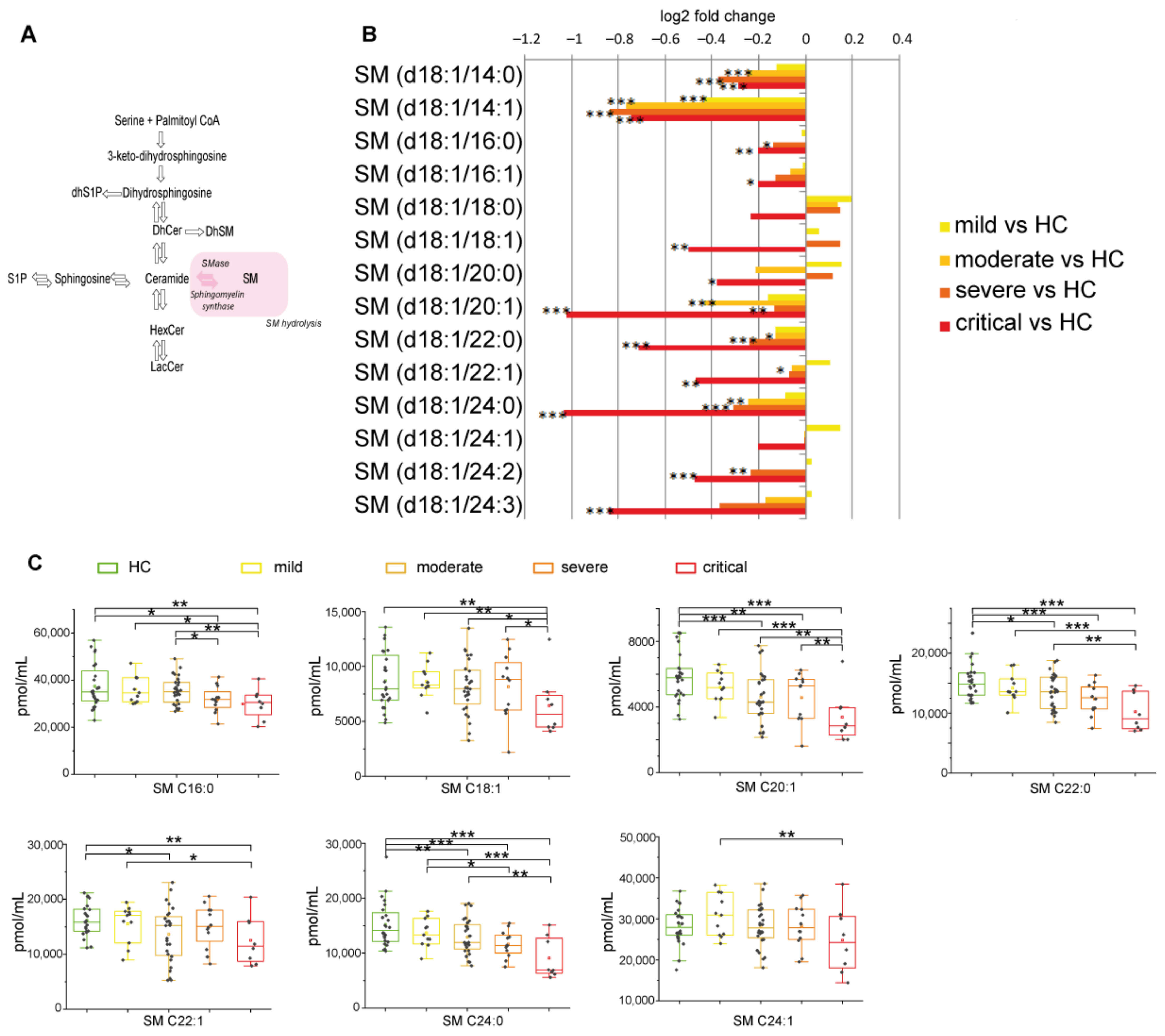
2.2.4. Sphingomyelin Decreased Reaching Lowest Levels in Severe and Critical Patients
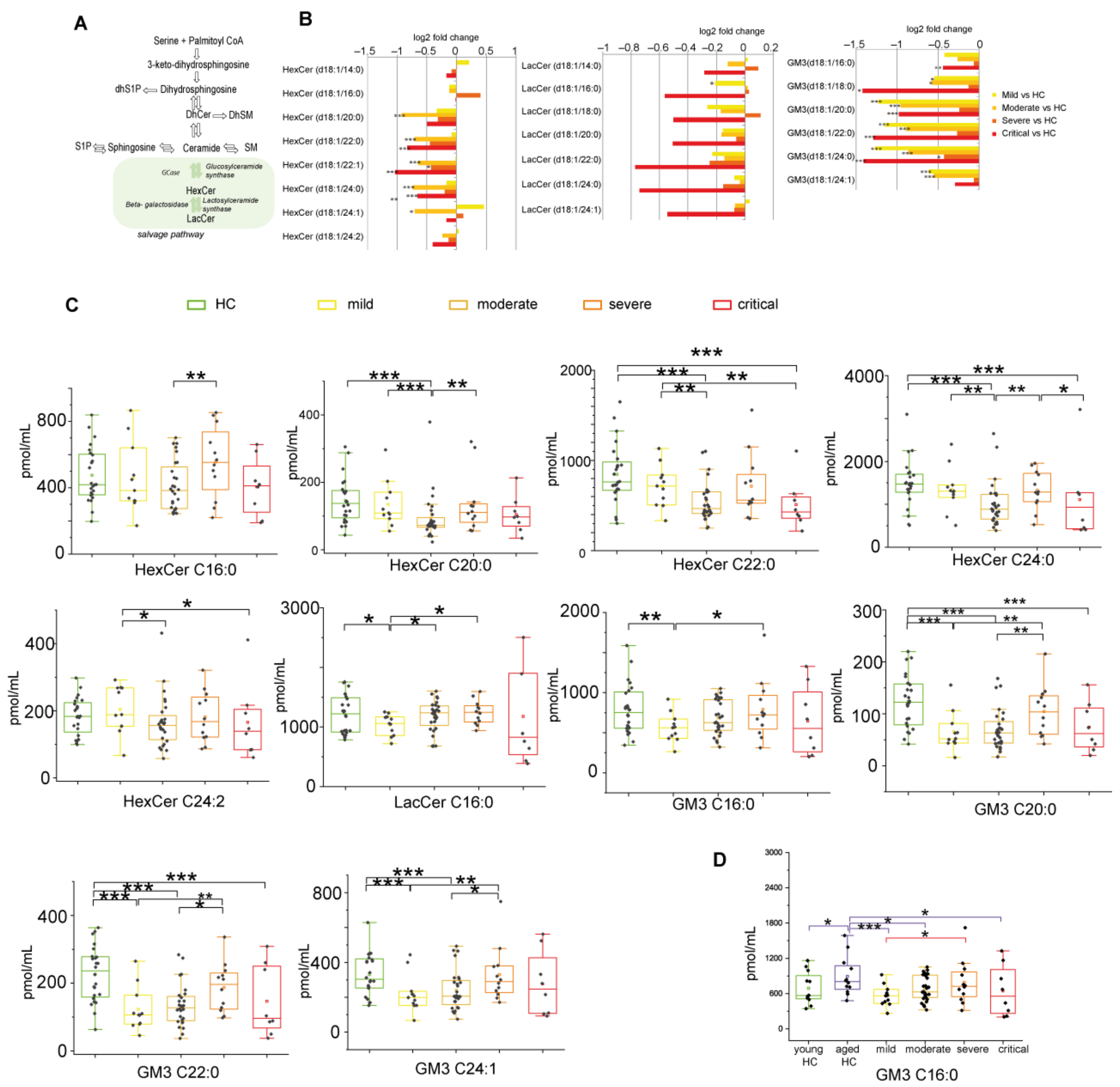
2.2.5. Glycosphingolipids and Disease Severity in COVID-19
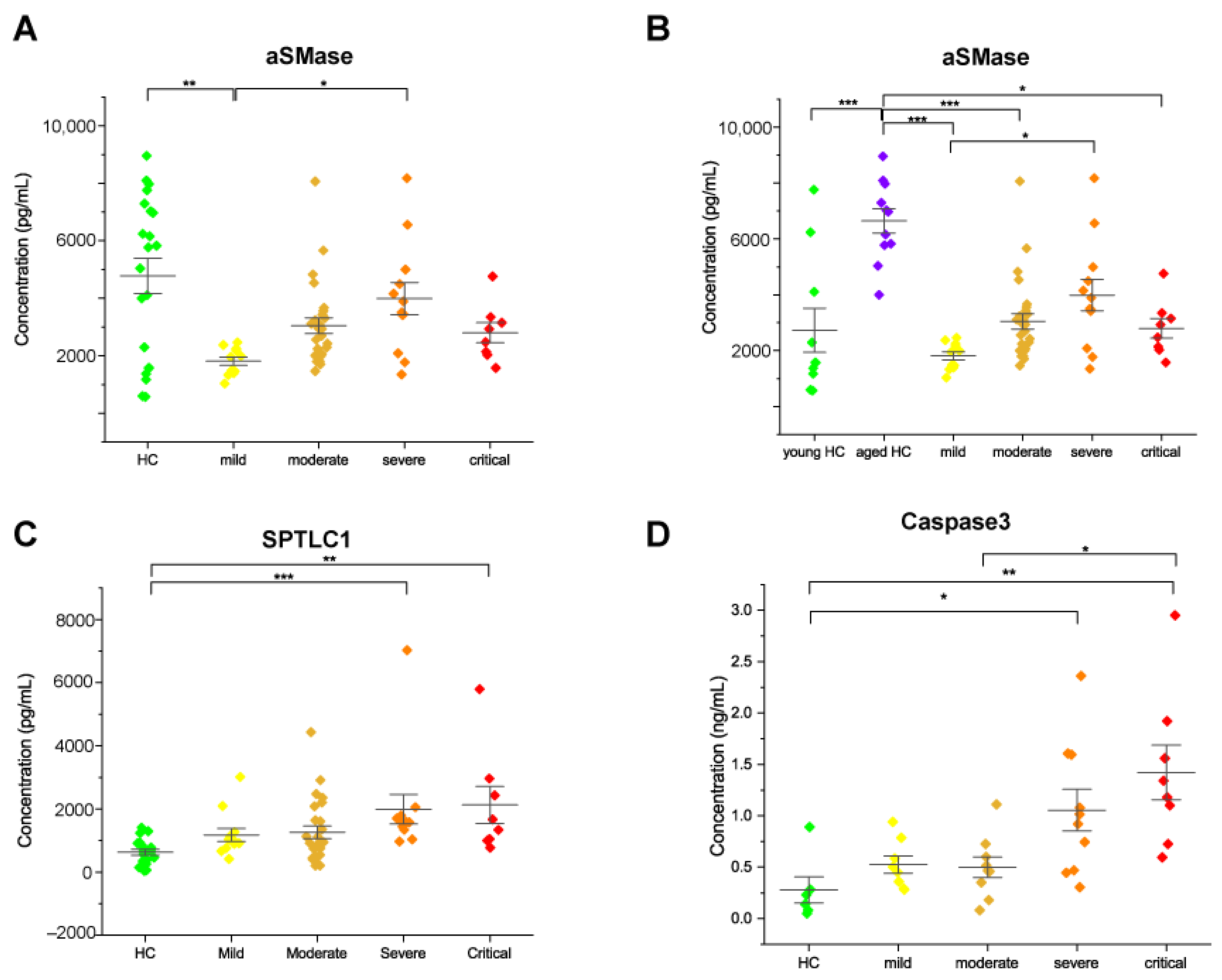
2.3. Acid Sphingomyelinase Levels Increased with COVID-19 Severity
2.4. Serine Palmitoyltransferase and Caspase 3 Levels Increased in Severe and Critical Patients
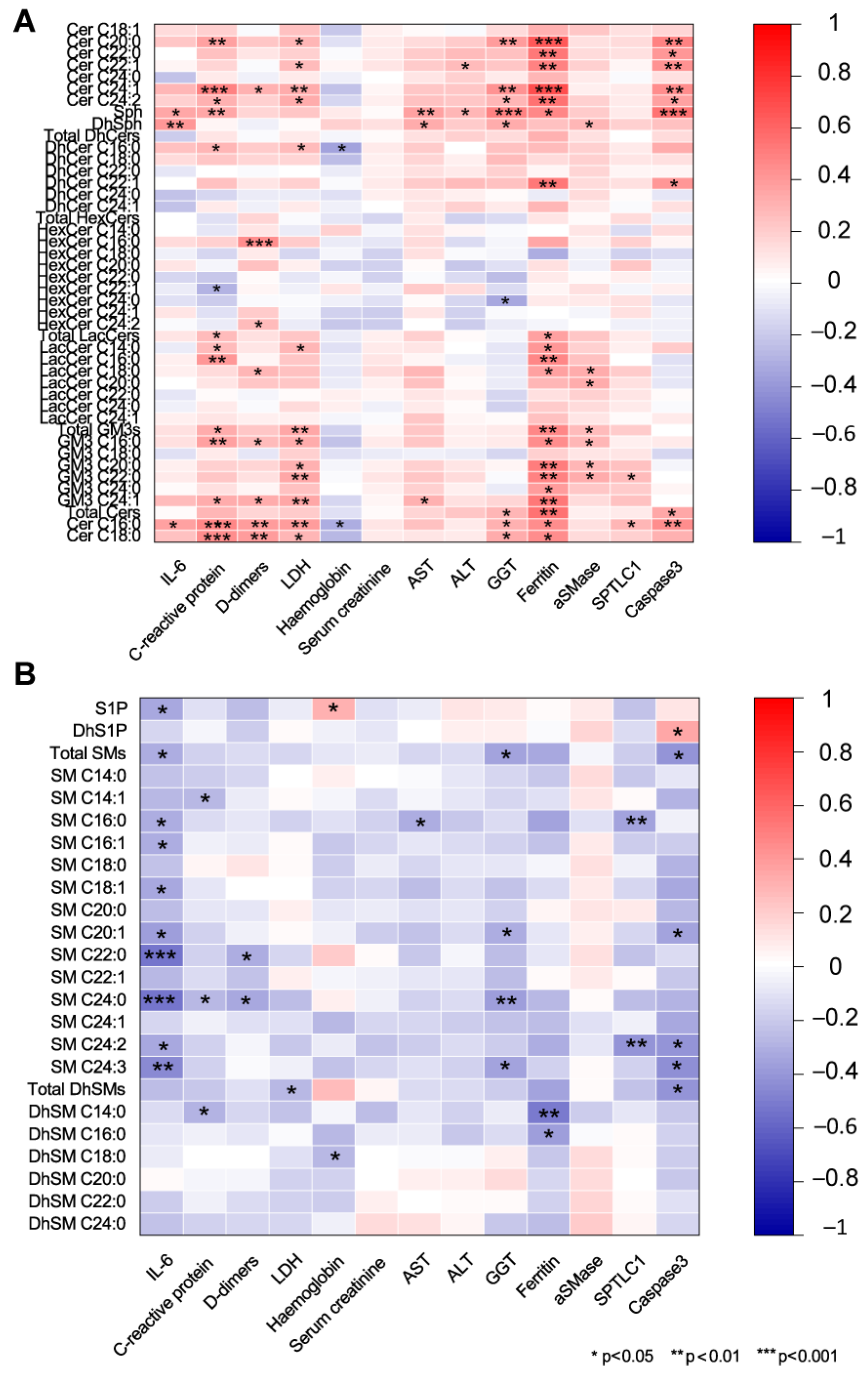
2.5. Association of Serum Sphingolipids with Biochemical Parameters in COVID-19
2.6. Sphingolipids as Biomarkers for COVID-19 Disease
3. Discussion
4. Materials and Methods
4.1. Participants and Sample Collection
4.2. Reagents and Chemicals
4.3. Lipid Extraction
4.4. Untargeted Lipidomics
4.5. Targeted Lipidomics
4.6. Enzyme-Linked Immunosorbent Assays
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gandhi, R.T.; Lynch, J.B.; Del Rio, C. Mild or Moderate Covid-19. N. Engl. J. Med. 2020, 383, 1757–1766. [Google Scholar] [CrossRef] [PubMed]
- Murthy, S.; Gomersall, C.D.; Fowler, R.A. Care for Critically Ill Patients With COVID-19. JAMA 2020, 323, 1499–1500. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; McGoogan, J.M. Characteristics of and Important Lessons From the Coronavirus Disease 2019 (COVID-19) Outbreak in China: Summary of a Report of 72314 Cases From the Chinese Center for Disease Control and Prevention. JAMA 2020, 323, 1239–1242. [Google Scholar] [CrossRef]
- Wang, H.; Yang, P.; Liu, K.; Guo, F.; Zhang, Y.; Zhang, G.; Jiang, C. SARS coronavirus entry into host cells through a novel clathrin- and caveolae-independent endocytic pathway. Cell Res. 2008, 18, 290–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeager, C.L.; Ashmun, R.A.; Williams, R.K.; Cardellichio, C.B.; Shapiro, L.H.; Look, A.T.; Holmes, K.V. Human aminopeptidase N is a receptor for human coronavirus 229E. Nature 1992, 357, 420–422. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Moore, M.J.; Vasilieva, N.; Sui, J.; Wong, S.K.; Berne, M.A.; Somasundaran, M.; Sullivan, J.L.; Luzuriaga, K.; Greenough, T.C.; et al. Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus. Nature 2003, 426, 450–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Deng, W.; Li, S.; Yang, X. Advances in research on ACE2 as a receptor for 2019-nCoV. Cell. Mol. Life Sci. CMLS 2021, 78, 531–544. [Google Scholar] [CrossRef] [PubMed]
- Peacock, T.P.; Goldhill, D.H.; Zhou, J.; Baillon, L.; Frise, R.; Swann, O.C.; Kugathasan, R.; Penn, R.; Brown, J.C.; Sanchez-David, R.Y.; et al. The furin cleavage site in the SARS-CoV-2 spike protein is required for transmission in ferrets. Nat. Microbiol. 2021, 6, 899–909. [Google Scholar] [CrossRef]
- Sviridov, D.; Miller, Y.I.; Ballout, R.A.; Remaley, A.T.; Bukrinsky, M. Targeting Lipid Rafts-A Potential Therapy for COVID-19. Front. Immunol. 2020, 11, 574508. [Google Scholar] [CrossRef]
- Fecchi, K.; Anticoli, S.; Peruzzu, D.; Iessi, E.; Gagliardi, M.C.; Matarrese, P.; Ruggieri, A. Coronavirus Interplay With Lipid Rafts and Autophagy Unveils Promising Therapeutic Targets. Front. Microbiol. 2020, 11, 1821. [Google Scholar] [CrossRef]
- Reinke, S.N.; Gallart-Ayala, H.; Gomez, C.; Checa, A.; Fauland, A.; Naz, S.; Kamleh, M.A.; Djukanovic, R.; Hinks, T.S.; Wheelock, C.E. Metabolomics analysis identifies different metabotypes of asthma severity. Eur. Respir. J. 2017, 49, 1601740. [Google Scholar] [CrossRef] [Green Version]
- Guerrera, I.C.; Astarita, G.; Jais, J.P.; Sands, D.; Nowakowska, A.; Colas, J.; Sermet-Gaudelus, I.; Schuerenberg, M.; Piomelli, D.; Edelman, A.; et al. A novel lipidomic strategy reveals plasma phospholipid signatures associated with respiratory disease severity in cystic fibrosis patients. PLoS ONE 2009, 4, e7735. [Google Scholar] [CrossRef] [Green Version]
- Cruickshank-Quinn, C.I.; Jacobson, S.; Hughes, G.; Powell, R.L.; Petrache, I.; Kechris, K.; Bowler, R.; Reisdorph, N. Metabolomics and transcriptomics pathway approach reveals outcome-specific perturbations in COPD. Sci. Rep. 2018, 8, 17132. [Google Scholar] [CrossRef]
- Tornquist, K.; Asghar, M.Y.; Srinivasan, V.; Korhonen, L.; Lindholm, D. Sphingolipids as Modulators of SARS-CoV-2 Infection. Front. Cell Dev. Biol. 2021, 9, 689854. [Google Scholar] [CrossRef]
- Schneider-Schaulies, J.; Schneider-Schaulies, S. Sphingolipids in viral infection. Biol. Chem. 2015, 396, 585–595. [Google Scholar] [CrossRef]
- Sakamoto, H.; Okamoto, K.; Aoki, M.; Kato, H.; Katsume, A.; Ohta, A.; Tsukuda, T.; Shimma, N.; Aoki, Y.; Arisawa, M.; et al. Host sphingolipid biosynthesis as a target for hepatitis C virus therapy. Nat. Chem. Biol. 2005, 1, 333–337. [Google Scholar] [CrossRef]
- Nguyen, A.; Guedan, A.; Mousnier, A.; Swieboda, D.; Zhang, Q.; Horkai, D.; Le Novere, N.; Solari, R.; Wakelam, M.J.O. Host lipidome analysis during rhinovirus replication in HBECs identifies potential therapeutic targets. J. Lipid Res. 2018, 59, 1671–1684. [Google Scholar] [CrossRef] [Green Version]
- Leier, H.C.; Weinstein, J.B.; Kyle, J.E.; Lee, J.Y.; Bramer, L.M.; Stratton, K.G.; Kempthorne, D.; Navratil, A.R.; Tafesse, E.G.; Hornemann, T.; et al. A global lipid map defines a network essential for Zika virus replication. Nat. Commun. 2020, 11, 3652. [Google Scholar] [CrossRef]
- Grammatikos, G.; Dietz, J.; Ferreiros, N.; Koch, A.; Dultz, G.; Bon, D.; Karakasiliotis, I.; Lutz, T.; Knecht, G.; Gute, P.; et al. Persistence of HCV in Acutely-Infected Patients Depletes C24-Ceramide and Upregulates Sphingosine and Sphinganine Serum Levels. Int. J. Mol. Sci. 2016, 17, 922. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.; Wang, X.; Deik, A.A.; Hanna, D.B.; Wang, T.; Haberlen, S.A.; Shah, S.J.; Lazar, J.M.; Hodis, H.N.; Landay, A.L.; et al. Elevated Plasma Ceramides Are Associated With Antiretroviral Therapy Use and Progression of Carotid Artery Atherosclerosis in HIV Infection. Circulation 2019, 139, 2003–2011. [Google Scholar] [CrossRef]
- Qu, F.; Zheng, S.J.; Liu, S.; Wu, C.S.; Duan, Z.P.; Zhang, J.L. Serum sphingolipids reflect the severity of chronic HBV infection and predict the mortality of HBV-acute-on-chronic liver failure. PLoS ONE 2014, 9, e104988. [Google Scholar] [CrossRef] [Green Version]
- Carpinteiro, A.; Edwards, M.J.; Hoffmann, M.; Kochs, G.; Gripp, B.; Weigang, S.; Adams, C.; Carpinteiro, E.; Gulbins, A.; Keitsch, S.; et al. Pharmacological Inhibition of Acid Sphingomyelinase Prevents Uptake of SARS-CoV-2 by Epithelial Cells. Cell Rep. Med. 2020, 1, 100142. [Google Scholar] [CrossRef] [PubMed]
- Carpinteiro, A.; Gripp, B.; Hoffmann, M.; Pohlmann, S.; Hoertel, N.; Edwards, M.J.; Kamler, M.; Kornhuber, J.; Becker, K.A.; Gulbins, E. Inhibition of acid sphingomyelinase by ambroxol prevents SARS-CoV-2 entry into epithelial cells. J. Biol. Chem. 2021, 296, 100701. [Google Scholar] [CrossRef]
- Hammad, S.M.; Pierce, J.S.; Soodavar, F.; Smith, K.J.; Al Gadban, M.M.; Rembiesa, B.; Klein, R.L.; Hannun, Y.A.; Bielawski, J.; Bielawska, A. Blood sphingolipidomics in healthy humans: Impact of sample collection methodology. J. Lipid Res. 2010, 51, 3074–3087. [Google Scholar] [CrossRef] [Green Version]
- Song, J.W.; Lam, S.M.; Fan, X.; Cao, W.J.; Wang, S.Y.; Tian, H.; Chua, G.H.; Zhang, C.; Meng, F.P.; Xu, Z.; et al. Omics-Driven Systems Interrogation of Metabolic Dysregulation in COVID-19 Pathogenesis. Cell Metab. 2020, 32, 188–202.e5. [Google Scholar] [CrossRef]
- Cuvillier, O.; Pirianov, G.; Kleuser, B.; Vanek, P.G.; Coso, O.A.; Gutkind, S.; Spiegel, S. Suppression of ceramide-mediated programmed cell death by sphingosine-1-phosphate. Nature 1996, 381, 800–803. [Google Scholar] [CrossRef]
- Spiegel, S.; Milstien, S. Sphingosine-1-phosphate: An enigmatic signalling lipid. Nat. Rev. Mol. Cell Biol. 2003, 4, 397–407. [Google Scholar] [CrossRef]
- Marfia, G.; Navone, S.; Guarnaccia, L.; Campanella, R.; Mondoni, M.; Locatelli, M.; Barassi, A.; Fontana, L.; Palumbo, F.; Garzia, E.; et al. Decreased serum level of sphingosine-1-phosphate: A novel predictor of clinical severity in COVID-19. EMBO Mol. Med. 2021, 13, e13424. [Google Scholar] [CrossRef] [PubMed]
- Van Echten-Deckert, G.; Zschoche, A.; Bar, T.; Schmidt, R.R.; Raths, A.; Heinemann, T.; Sandhoff, K. cis-4-Methylsphingosine decreases sphingolipid biosynthesis by specifically interfering with serine palmitoyltransferase activity in primary cultured neurons. J. Biol. Chem. 1997, 272, 15825–15833. [Google Scholar] [CrossRef] [Green Version]
- Katsume, A.; Tokunaga, Y.; Hirata, Y.; Munakata, T.; Saito, M.; Hayashi, H.; Okamoto, K.; Ohmori, Y.; Kusanagi, I.; Fujiwara, S.; et al. A serine palmitoyltransferase inhibitor blocks hepatitis C virus replication in human hepatocytes. Gastroenterology 2013, 145, 865–873. [Google Scholar] [CrossRef] [Green Version]
- Tippetts, T.S.; Holland, W.L.; Summers, S.A. The ceramide ratio: A predictor of cardiometabolic risk. J. Lipid Res. 2018, 59, 1549–1550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lingwood, D.; Simons, K. Lipid rafts as a membrane-organizing principle. Science 2010, 327, 46–50. [Google Scholar] [CrossRef] [Green Version]
- Patschan, S.; Chen, J.; Gealekman, O.; Krupincza, K.; Wang, M.; Shu, L.; Shayman, J.A.; Goligorsky, M.S. Mapping mechanisms and charting the time course of premature cell senescence and apoptosis: Lysosomal dysfunction and ganglioside accumulation in endothelial cells. Am. J. Physiol. Ren. Physiol. 2008, 294, F100–F109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennaceur, K.; Popa, I.; Portoukalian, J.; Berthier-Vergnes, O.; Peguet-Navarro, J. Melanoma-derived gangliosides impair migratory and antigen-presenting function of human epidermal Langerhans cells and induce their apoptosis. Int. Immunol. 2006, 18, 879–886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennaceur, K.; Popa, I.; Chapman, J.A.; Migdal, C.; Peguet-Navarro, J.; Touraine, J.L.; Portoukalian, J. Different mechanisms are involved in apoptosis induced by melanoma gangliosides on human monocyte-derived dendritic cells. Glycobiology 2009, 19, 576–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torretta, E.; Barbacini, P.; Al-Daghri, N.M.; Gelfi, C. Sphingolipids in Obesity and Correlated Co-Morbidities: The Contribution of Gender, Age and Environment. Int. J. Mol. Sci. 2019, 20, 5901. [Google Scholar] [CrossRef] [Green Version]
- Al-Daghri, N.M.; Torretta, E.; Barbacini, P.; Asare, H.; Ricci, C.; Capitanio, D.; Guerini, F.R.; Sabico, S.B.; Alokail, M.S.; Clerici, M.; et al. Sphingolipid serum profiling in vitamin D deficient and dyslipidemic obese dimorphic adults. Sci. Rep. 2019, 9, 16664. [Google Scholar] [CrossRef]
- Barbacini, P.; Casas, J.; Torretta, E.; Capitanio, D.; Maccallini, G.; Hirschler, V.; Gelfi, C. Regulation of Serum Sphingolipids in Andean Children Born and Living at High Altitude (3775 m). Int. J. Mol. Sci. 2019, 20, 2835. [Google Scholar] [CrossRef] [Green Version]
- Poss, A.M.; Maschek, J.A.; Cox, J.E.; Hauner, B.J.; Hopkins, P.N.; Hunt, S.C.; Holland, W.L.; Summers, S.A.; Playdon, M.C. Machine learning reveals serum sphingolipids as cholesterol-independent biomarkers of coronary artery disease. J. Clin. Investig. 2020, 130, 1363–1376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montoliu, I.; Scherer, M.; Beguelin, F.; DaSilva, L.; Mari, D.; Salvioli, S.; Martin, F.P.; Capri, M.; Bucci, L.; Ostan, R.; et al. Serum profiling of healthy aging identifies phospho- and sphingolipid species as markers of human longevity. Aging 2014, 6, 9–25. [Google Scholar] [CrossRef] [Green Version]
- Torretta, E.; Arosio, B.; Barbacini, P.; Casati, M.; Capitanio, D.; Mancuso, R.; Mari, D.; Cesari, M.; Clerici, M.; Gelfi, C. Particular CSF sphingolipid patterns identify iNPH and AD patients. Sci. Rep. 2018, 8, 13639. [Google Scholar] [CrossRef]
- Silva, L.; de Almeida, R.F.; Fedorov, A.; Matos, A.P.; Prieto, M. Ceramide-platform formation and -induced biophysical changes in a fluid phospholipid membrane. Mol. Membr. Biol. 2006, 23, 137–148. [Google Scholar] [CrossRef]
- Kolesnick, R.N.; Goni, F.M.; Alonso, A. Compartmentalization of ceramide signaling: Physical foundations and biological effects. J. Cell. Physiol. 2000, 184, 285–300. [Google Scholar] [CrossRef]
- Slotte, J.P.; Bierman, E.L. Movement of plasma-membrane sterols to the endoplasmic reticulum in cultured cells. Biochem. J. 1987, 248, 237–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zha, X.; Pierini, L.M.; Leopold, P.L.; Skiba, P.J.; Tabas, I.; Maxfield, F.R. Sphingomyelinase treatment induces ATP-independent endocytosis. J. Cell Biol. 1998, 140, 39–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edwards, M.J.; Becker, K.A.; Gripp, B.; Hoffmann, M.; Keitsch, S.; Wilker, B.; Soddemann, M.; Gulbins, A.; Carpinteiro, E.; Patel, S.H.; et al. Sphingosine prevents binding of SARS-CoV-2 spike to its cellular receptor ACE2. J. Biol. Chem. 2020, 295, 15174–15182. [Google Scholar] [CrossRef]
- Zhou, C.; Chen, Y.; Ji, Y.; He, X.; Xue, D. Increased Serum Levels of Hepcidin and Ferritin Are Associated with Severity of COVID-19. Med Sci. Monit. Int. Med J. Exp. Clin. Res. 2020, 26, e926178. [Google Scholar]
- Ottolenghi, S.; Zulueta, A.; Caretti, A. Iron and Sphingolipids as Common Players of (Mal)Adaptation to Hypoxia in Pulmonary Diseases. Int. J. Mol. Sci. 2020, 21, 307. [Google Scholar] [CrossRef] [Green Version]
- Janneh, A.H.; Kassir, M.F.; Dwyer, C.J.; Chakraborty, P.; Pierce, J.S.; Flume, P.A.; Li, H.; Nadig, S.N.; Mehrotra, S.; Ogretmen, B. Alterations of lipid metabolism provide serologic biomarkers for the detection of asymptomatic versus symptomatic COVID-19 patients. Sci. Rep. 2021, 11, 14232. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Healthy Controls (n = 24) | Mild (n = 11) | Moderate (n = 28) | Severe (n = 12) | Critical (n = 8) | ||
|---|---|---|---|---|---|---|
| Characteristics | p-value | |||||
| Age, years | 52 (26–78) | 45 (19–85) | 55 (24–98) | 62.5 (40–84) | 63.5 (49–75) | 0.276 |
| Sex | 0.194 | |||||
| Men | 11 (46%) | 3 (27%) | 16 (57%) | 8 (67%) | 6 (75%) | |
| Women | 13 (54%) | 8(73%) | 12 (43%) | 4 (33%) | 2 (25%) | |
| Co-infections | ||||||
| Hepatitis B | 0 | 1 (9%) | 0 | 0 | 0 | |
| Syphilis | 0 | 0 | 1 (3%) | 0 | 0 | |
| Co-morbidities | ||||||
| Hypertension | 5 (45%) | 9 (32%) | 6 (50%) | 4 (50%) | 0.643 | |
| Diabetes | 2 (18%) | 5 (18%) | 2 (17%) | 0 | 0.969 | |
| Dyslipidemia | 2 (18%) | 1 (4%) | 5 (42%) | 0 | ||
| Obesity | 0 | 3 (11%) | 1 (8%) | 0 | ||
| Solid/hematologic tumors | 0 | 1 (4%) | 3 (25%) | 0 | ||
| COPD | 0 | 1 (4%) | 1 (8%) | 1 (12%) | ||
| Death | 0 | 1 (9%) | 0 | 1 (8%) | 2 (25%) | |
| Haemoglobin, g/dL | 12.6 (10.3–17.2) | 14.2 (8.7–17.5) | 12.5 (10.2–14.2) | 10.3 (8.1–14.4) | 0.024 | |
| Platelets, ×103 cells/mm3 | 198 (145–416) | 209 (91–407) | 256 (98–661) | 251 (74–483) | 0.66 | |
| White Blood Cells, ×103 cells/mm3 | 3800 (1790–6650) | 5390 (1460–9940) | 6095 (1580–11,150) | 6380 (3950–42,510) | 0.0265 | |
| Neutrophils, ×103 cells/mm3 | 3.745 (1.26–6.55) | 3.14 (1.46–21.67) | 4.265 (1.77–6.37) | 4.97 (2.04–23.55) | 0.261 | |
| Lymphocytes, ×103 cells/mm3 | 1.15 (0.43–1.96) | 1.165 (0.3–221) | 0.79 (0.45–1.71) | 1.07 (0.5–2.19) | 0.281 | |
| Interleukin-6, pg/mL | 3.21 (0.44–26.32) | 10.615 (1.83–70.59) | 120.72 (36.11–215.41) | 126.27 (81.57–546.78) | <0.001 | |
| C-reactive protein, mg/dL | 9.3 (0.3–86.1) | 40.05 (0.8–320.1) | 66.4 (7.1–254) | 71.8 (1–176.7) | 0.0387 | |
| D-dimer, ng/mL | 488 (101–17,947) | 772.5 (218–36,910) | 1544 (421–5194) | 557 (369–5309) | 0.702 | |
| Lactate Dehydrogenase, UI/L | 166 (114–275) | 242 (54–941) | 251 (137–1424) | 361 (223–577) | 0.0016 | |
| Serum creatinine, mg/dL | 0.73 (0.58–1.43) | 0.85 (0.57–1.31) | 0.945 (0.42–4.39) | 0.895 (0.52–2.12) | 0.595 | |
| Aspartate Aminotransferase, mU/mL | 21 (14–54) | 22 (11–114) | 20 (0.99–234) | 43.5 (11–169) | 0.07318 | |
| Alanine Aminotransferase, mU/mL | 15 (9–66) | 25 (7–158) | 18 (8–103) | 41 (6–439) | 0.1933 | |
| Gamma-glutamyl transpeptidase, UI/L | 25 (12–84) | 27 (9–110) | 21 (10–354) | 97 (23–913) | 0.00977 | |
| Ferritin, ng/mL | 123 (11–378) | 206 (15–1000) | 647 (340–1000) | 886 (140–1000) | 0.01085 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Torretta, E.; Garziano, M.; Poliseno, M.; Capitanio, D.; Biasin, M.; Santantonio, T.A.; Clerici, M.; Lo Caputo, S.; Trabattoni, D.; Gelfi, C. Severity of COVID-19 Patients Predicted by Serum Sphingolipids Signature. Int. J. Mol. Sci. 2021, 22, 10198. https://doi.org/10.3390/ijms221910198
Torretta E, Garziano M, Poliseno M, Capitanio D, Biasin M, Santantonio TA, Clerici M, Lo Caputo S, Trabattoni D, Gelfi C. Severity of COVID-19 Patients Predicted by Serum Sphingolipids Signature. International Journal of Molecular Sciences. 2021; 22(19):10198. https://doi.org/10.3390/ijms221910198
Chicago/Turabian StyleTorretta, Enrica, Micaela Garziano, Mariacristina Poliseno, Daniele Capitanio, Mara Biasin, Teresa Antonia Santantonio, Mario Clerici, Sergio Lo Caputo, Daria Trabattoni, and Cecilia Gelfi. 2021. "Severity of COVID-19 Patients Predicted by Serum Sphingolipids Signature" International Journal of Molecular Sciences 22, no. 19: 10198. https://doi.org/10.3390/ijms221910198