
Protein Arginine Methyltransferase 6 Involved in Germ Cell Viability during Spermatogenesis and Down-Regulated by the Androgen Receptor

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
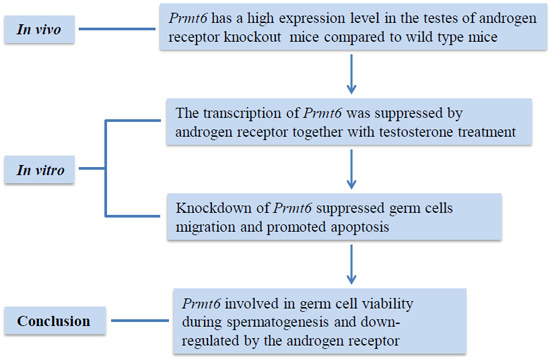
2. Results
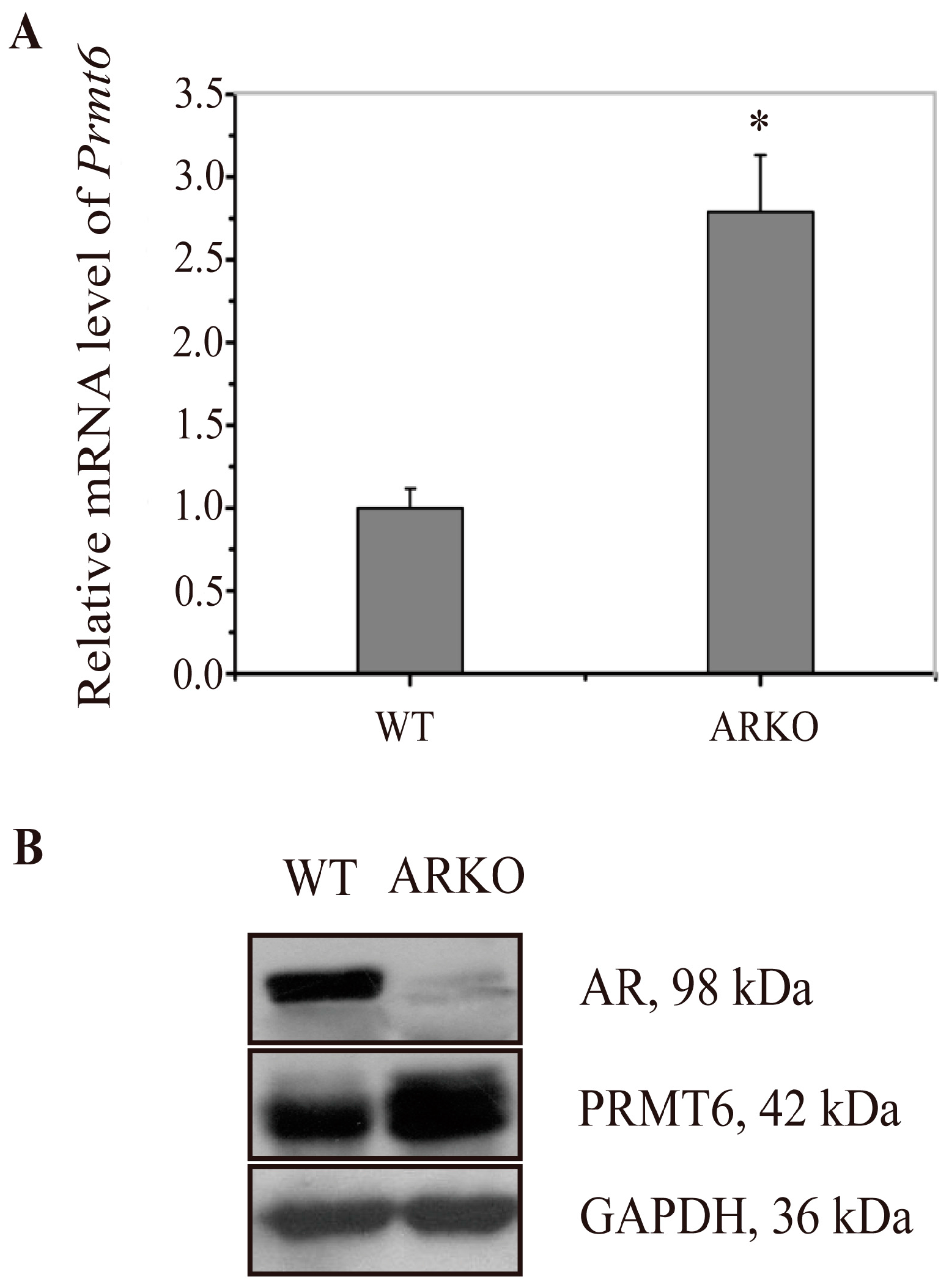
2.1. Prmt6 (Protein Arginine Methyltransferase 6) mRNA and PRMT6 Protein Expression Were Increased in the Testes of ARKO (Androgen Receptor Knockout) Mice

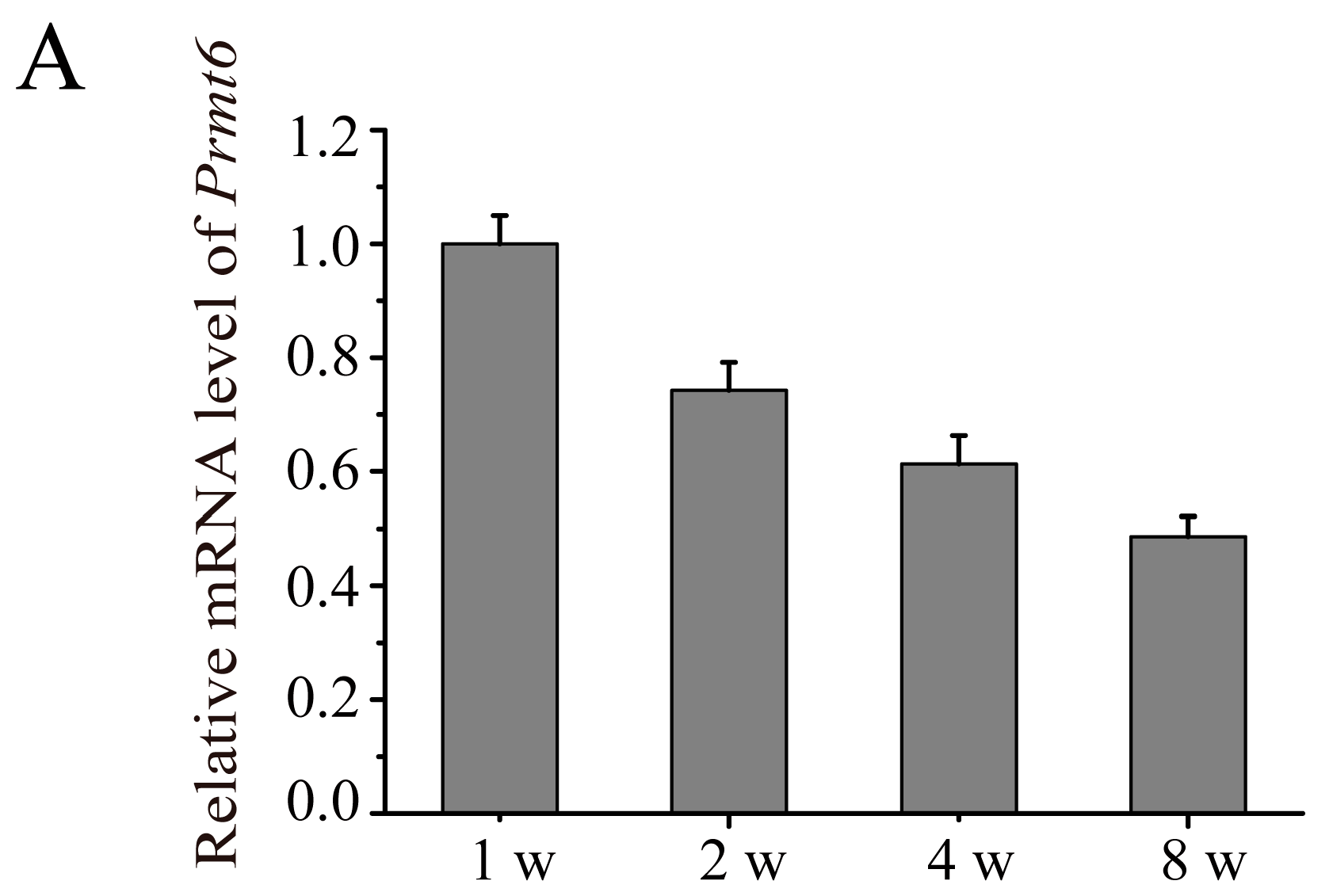
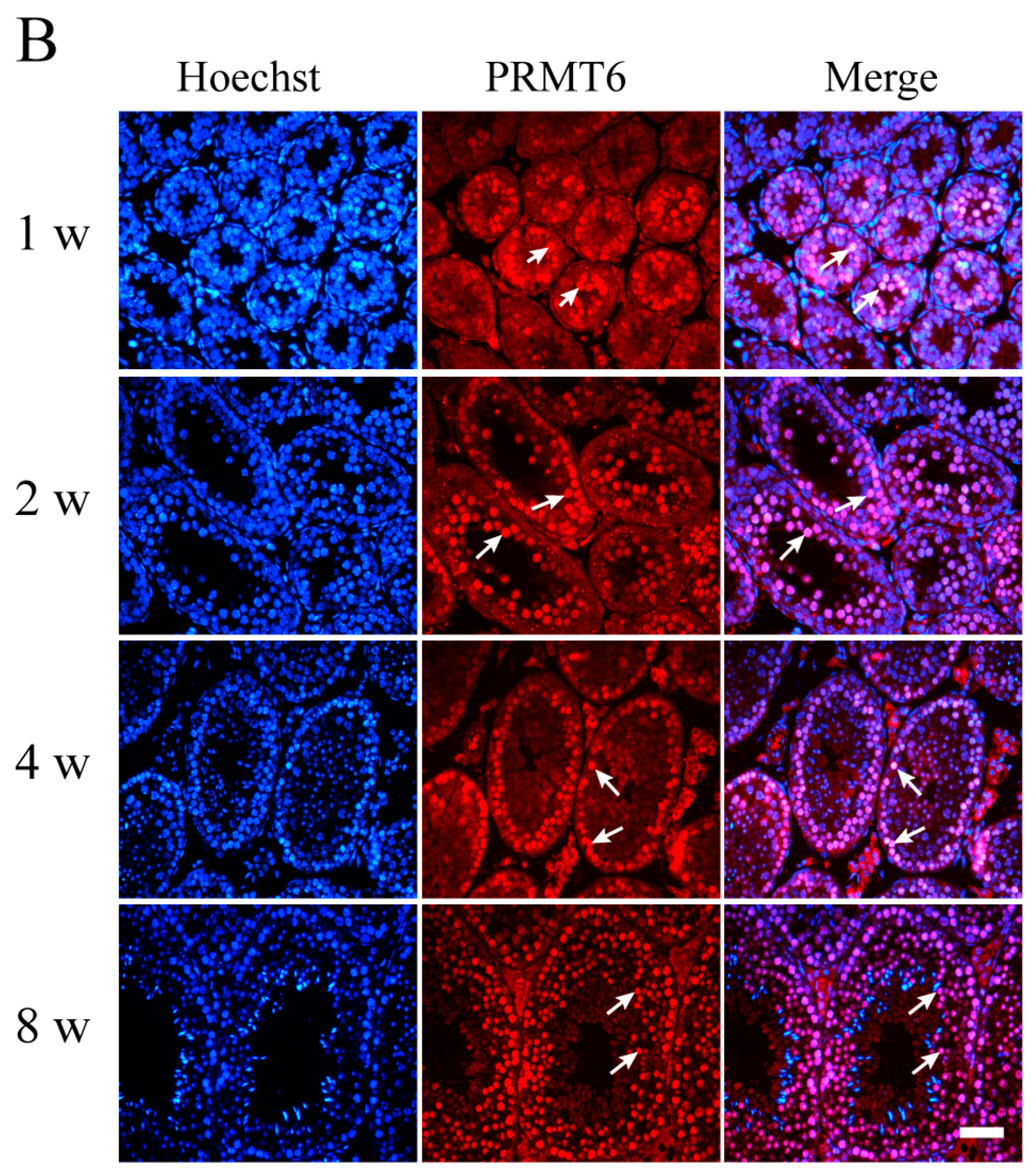
2.2. The Expression of Prmt6 mRNA and PRMT6 Protein Localization during Mouse Testes Development


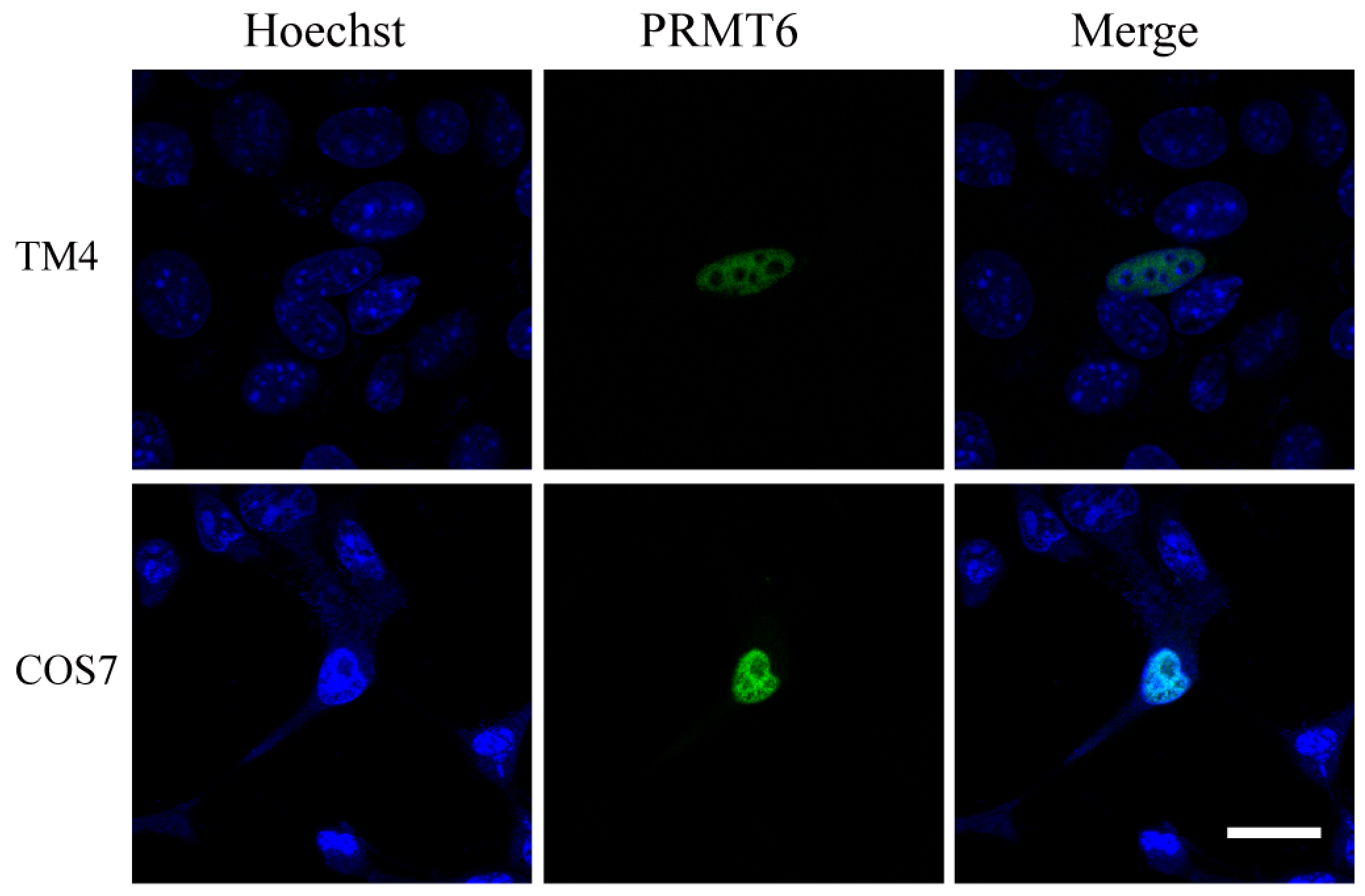
2.3. Sub-Cellular Localization of the EGFP (Enhanced Green Fluorescent Protein)-PRMT6 Fusion Protein

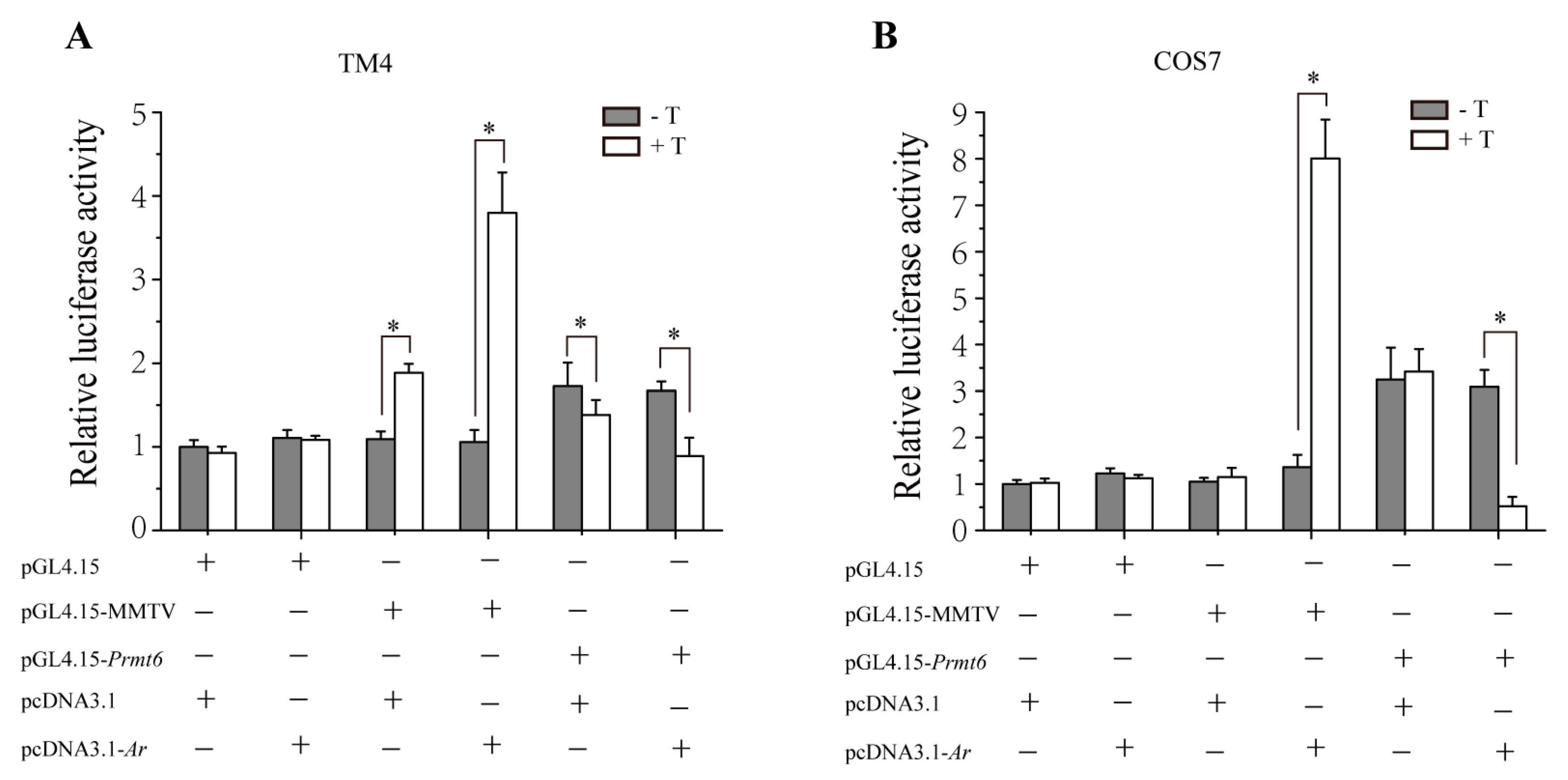
2.4. In Vitro Repression of Prmt6 Promoter Activity by Androgen Receptor (AR) and Testosterone

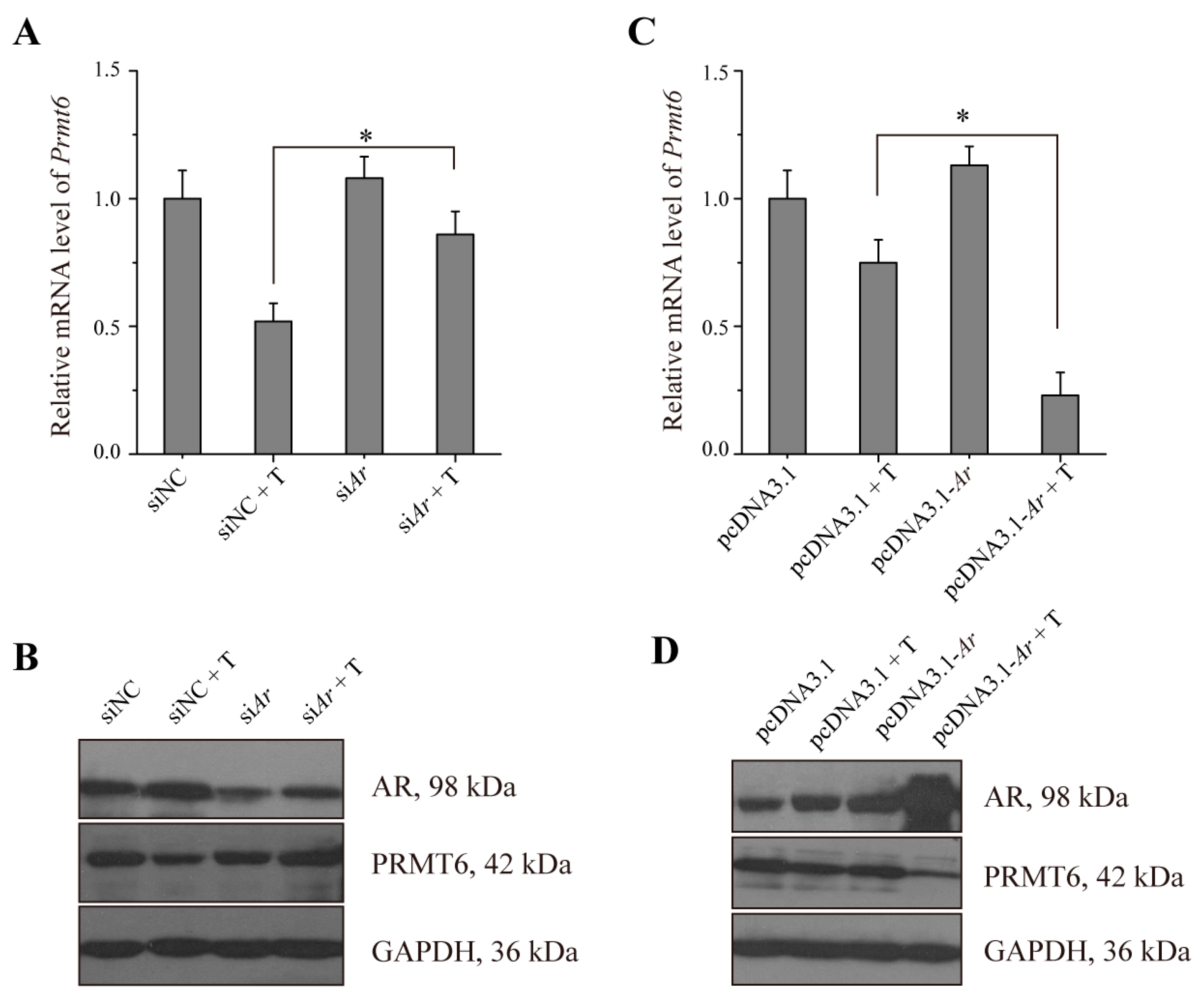
2.5. The Expression of Prmt6 mRNA and PRMT6 Protein Were Regulated by AR in Vitro

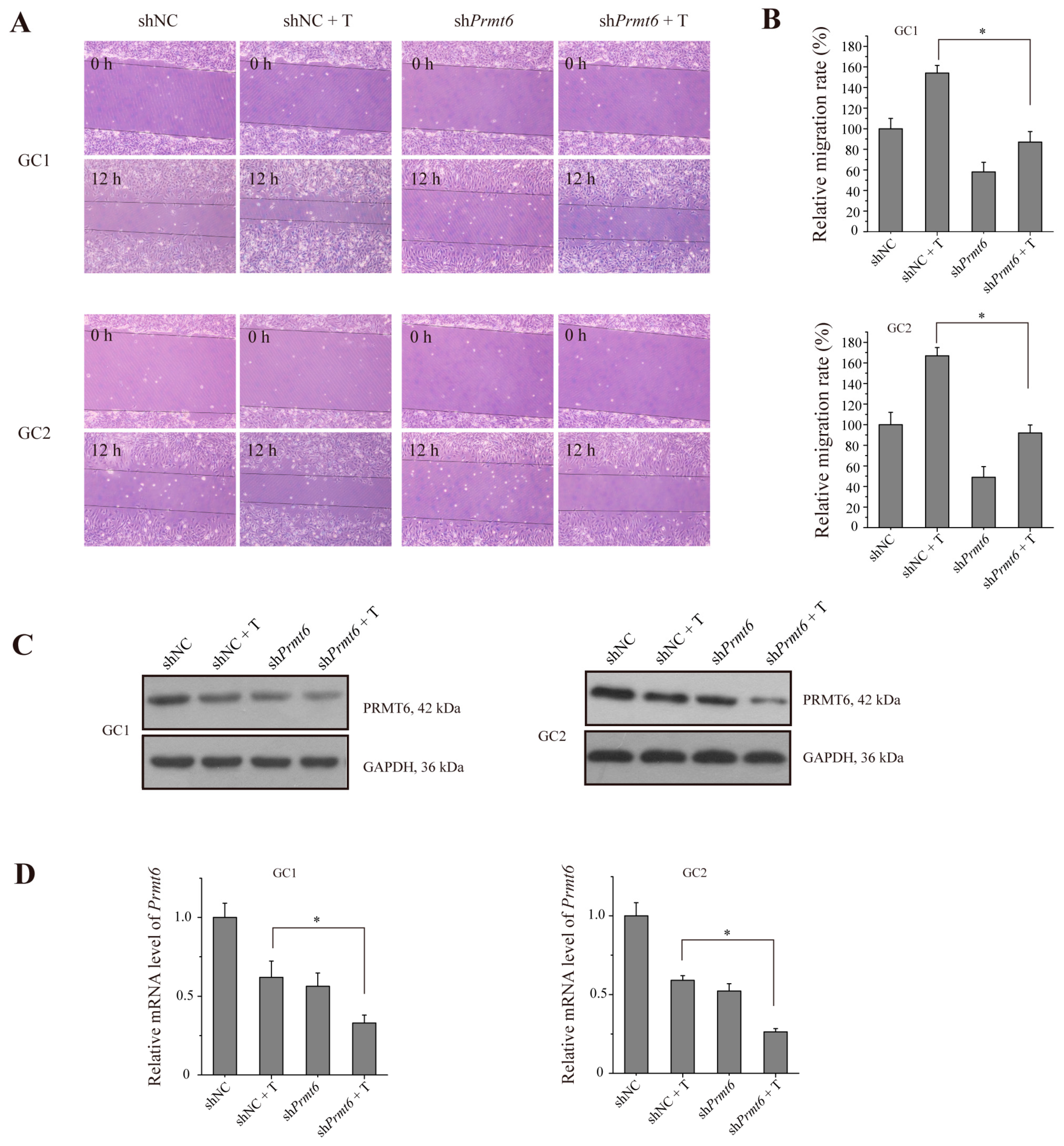
2.6. The Knockdown of Prmt6 Represses Germ Cell Migration

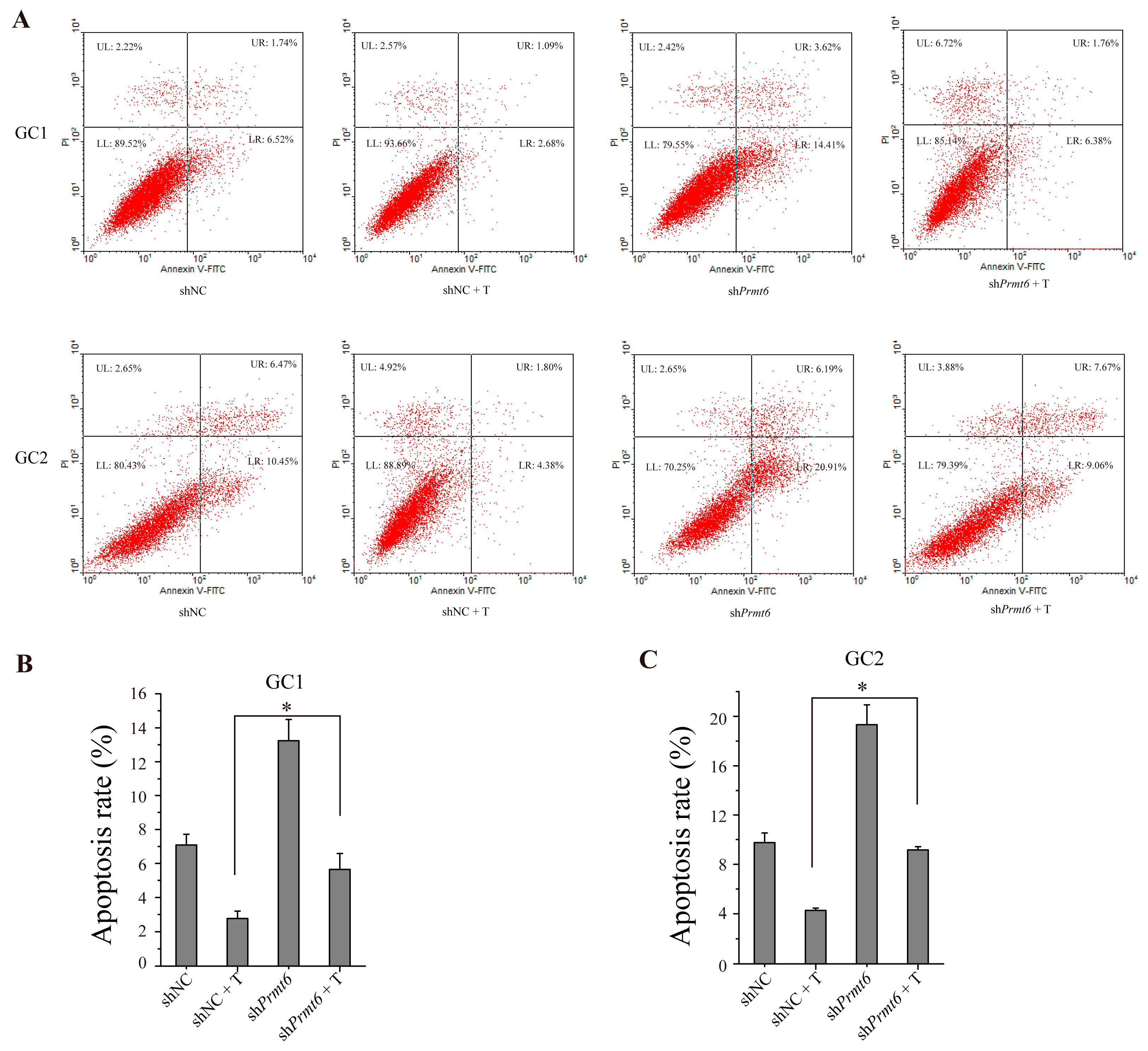
2.7. The Down-Regulation of Prmt6 Induces Germ Cell Apoptosis

3. Discussion
4. Experimental Section
4.1. Animals and Samples
4.2. Cell Culture
4.3. Total RNA Isolation and Quantitative Real-Time RT-PCR (RT-qPCR)
4.4. Western Blotting Analysis
4.5. Immunofluorescent Localization of PRMT6
4.6. Construction and Sub-Cellular Localization of EGFP-PRMT6 Fusion Vector
4.7. Plasmid Constructs and Luciferase Assay
4.8. RNA Interference
4.9. Short Hairpin RNA-Mediated Knockdown
4.10. Migration Assay
4.11. Flow Cytometry Analysis of Cell Apoptosis
4.12. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Heemers, H.V.; Tindall, D.J. Androgen receptor (AR) coregulators: A diversity of functions converging on and regulating the AR transcriptional complex. Endocr. Rev. 2007, 28, 778–808. [Google Scholar] [CrossRef] [PubMed]
- Patrao, M.T.; Silva, E.J.; Avellar, M.C. Androgens and the male reproductive tract: An overview of classical roles and current perspectives. Arq. Bras. Endocrinol. Metabol. 2009, 53, 934–945. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.S.; Yeh, S.; Tzeng, C.R.; Chang, C. Androgen receptor roles in spermatogenesis and fertility: Lessons from testicular cell-specific androgen receptor knockout mice. Endocr. Rev. 2009, 30, 119–132. [Google Scholar] [CrossRef] [PubMed]
- Heinlein, C.A.; Chang, C. Androgen receptor (AR) coregulators: An overview. Endocr. Rev. 2002, 23, 175–200. [Google Scholar] [CrossRef] [PubMed]
- Yeh, S.; Tsai, M.Y.; Xu, Q.; Mu, X.M.; Lardy, H.; Huang, K.E.; Lin, H.; Yeh, S.D.; Altuwaijri, S.; Zhou, X.; et al. Generation and characterization of androgen receptor knockout (ARKO) mice: An in vivo model for the study of androgen functions in selective tissues. Proc. Natl. Acad. Sci. USA 2002, 99, 13498–13503. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, A.; Rao, M.K.; Kaur, R.; Buttigieg, M.R.; Wilkinson, M.F. GATA factors and androgen receptor collaborate to transcriptionally activate the Rhox5 homeobox gene in sertoli cells. Mol. Cell. Boil. 2008, 28, 2138–2153. [Google Scholar] [CrossRef] [PubMed]
- De Gendt, K.; Denolet, E.; Willems, A.; Daniels, V.W.; Clinckemalie, L.; Denayer, S.; Wilkinson, M.F.; Claessens, F.; Swinnen, J.V.; Verhoeven, G. Expression of Tubb3, a β-tubulin isotype, is regulated by androgens in mouse and rat sertoli cells. Biol. Reprod. 2011, 85, 934–945. [Google Scholar] [CrossRef] [PubMed]
- Lim, K.; Yoo, J.H.; Kim, K.Y.; Kweon, G.R.; Kwak, S.T.; Hwang, B.D. Testosterone regulation of proto-oncogene c-myc expression in primary Sertoli cell cultures from prepubertal rats. J. Androl. 1994, 15, 543–550. [Google Scholar] [PubMed]
- Zhang, Q.X.; Zhang, X.Y.; Zhang, Z.M.; Lu, W.; Liu, L.; Li, G.; Cai, Z.M.; Gui, Y.T.; Chang, C. Identification of testosterone-/androgen receptor-regulated genes in mouse Sertoli cells. Asian J. Androl. 2012, 14, 294–300. [Google Scholar] [CrossRef] [PubMed]
- Meyer, R.; Wolf, S.S.; Obendorf, M. PRMT2, a member of the protein arginine methyltransferase family, is a coactivator of the androgen receptor. J. Steroid Biochem. Mol. Biol. 2007, 107, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Harada, N.; Takagi, T.; Nakano, Y.; Yamaji, R.; Inui, H. Protein arginine methyltransferase 10 is required for androgen-dependent proliferation of LNCaP prostate cancer cells. Biosci. Biotechnol. Biochem. 2015, 79, 1430–1437. [Google Scholar] [CrossRef] [PubMed]
- Scaramuzzino, C.; Casci, I.; Parodi, S.; Lievens, P.M.; Polanco, M.J.; Milioto, C.; Chivet, M.; Monaghan, J.; Mishra, A.; Badders, N.; et al. Protein arginine methyltransferase 6 enhances polyglutamine-expanded androgen receptor function and toxicity in spinal and bulbar muscular atrophy. Neuron 2015, 85, 88–100. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Chung, H.H.; Woo, A.R.; Lin, V.C. Protein arginine methyltransferase 6 enhances ligand-dependent and -independent activity of estrogen receptor α via distinct mechanisms. Biochim. Biophys. Acta 2014, 1843, 2067–2078. [Google Scholar] [CrossRef] [PubMed]
- Tu, W.; Liu, Y.; Shen, Y.; Yan, Y.; Wang, X.; Yang, D.; Li, L.; Ma, Y.; Tao, D.; Zhang, S.; et al. Genome-wide loci linked to non-obstructive azoospermia susceptibility may be independent of reduced sperm production in males with normozoospermia. Biol. Reprod. 2015, 92, 41. [Google Scholar] [CrossRef] [PubMed]
- Frankel, A.; Yadav, N.; Lee, J.; Branscombe, T.L.; Clarke, S.; Bedford, M.T. The novel human protein arginine N-methyltransferase PRMT6 is a nuclear enzyme displaying unique substrate specificity. J. Biol. Chem. 2002, 277, 3537–3543. [Google Scholar] [CrossRef] [PubMed]
- Guccione, E.; Bassi, C.; Casadio, F.; Martinato, F.; Cesaroni, M.; Schuchlautz, H.; Luscher, B.; Amati, B. Methylation of histone H3R2 by PRMT6 and H3K4 by an MLL complex are mutually exclusive. Nature 2007, 449, 933–937. [Google Scholar] [CrossRef] [PubMed]
- Hyllus, D.; Stein, C.; Schnabel, K.; Schiltz, E.; Imhof, A.; Dou, Y.; Hsieh, J.; Bauer, U.M. PRMT6-mediated methylation of R2 in histone H3 antagonizes H3K4 trimethylation. Genes Dev. 2007, 21, 3369–3380. [Google Scholar] [CrossRef] [PubMed]
- Harrison, M.J.; Tang, Y.H.; Dowhan, D.H. Protein arginine methyltransferase 6 regulates multiple aspects of gene expression. Nucleic Acids Res. 2010, 38, 2201–2216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michaud-Levesque, J.; Richard, S. Thrombospondin-1 is a transcriptional repression target of PRMT6. J. Biol. Chem. 2009, 284, 21338–21346. [Google Scholar] [CrossRef] [PubMed]
- Phalke, S.; Mzoughi, S.; Bezzi, M.; Jennifer, N.; Mok, W.C.; Low, D.H.; Thike, A.A.; Kuznetsov, V.A.; Tan, P.H.; Voorhoeve, P.M.; et al. P53-independent regulation of p21Waf1/Cip1 expression and senescence by PRMT6. Nucleic Acids Res. 2012, 40, 9534–9542. [Google Scholar] [CrossRef] [PubMed]
- El-Andaloussi, N.; Valovka, T.; Toueille, M.; Steinacher, R.; Focke, F.; Gehrig, P.; Covic, M.; Hassa, P.O.; Schar, P.; Hubscher, U.; et al. Arginine methylation regulates DNA polymerase β. Mol. Cell 2006, 22, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Olsen, A.K.; Bjortuft, H.; Wiger, R.; Holme, J.; Seeberg, E.; Bjoras, M.; Brunborg, G. Highly efficient base excision repair (BER) in human and rat male germ cells. Nucleic Acids Res. 2001, 29, 1781–1790. [Google Scholar] [CrossRef] [PubMed]
- Plug, A.W.; Clairmont, C.A.; Sapi, E.; Ashley, T.; Sweasy, J.B. Evidence for a role for DNA polymerase β in mammalian meiosis. Proc. Natl Acad. Sci. USA 1997, 94, 1327–1331. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Xia, Y.; Guo, X.; Dai, J.; Li, H.; Hu, H.; Jiang, Y.; Lu, F.; Wu, Y.; Yang, X.; et al. A genome-wide association study in Chinese men identifies three risk loci for non-obstructive azoospermia. Nat. Genet. 2012, 44, 183–186. [Google Scholar] [CrossRef] [PubMed]
- Mirfakhraie, R.; Kalantar, S.M.; Mirzajani, F.; Montazeri, M.; Salsabili, N.; Houshmand, M.; Hashemi-Gorji, F.; Pourmand, G. A novel mutation in the transactivation-regulating domain of the androgen receptor in a patient with azoospermia. J. Androl. 2011, 32, 367–370. [Google Scholar] [CrossRef] [PubMed]
- Goglia, U.; Vinanzi, C.; Zuccarello, D.; Malpassi, D.; Ameri, P.; Casu, M.; Minuto, F.; Foresta, C.; Ferone, D. Identification of a novel mutation in exon 1 of androgen receptor gene in an azoospermic patient with mild androgen insensitivity syndrome: Case report and literature review. Fertil. Steril. 2011, 96, 1165–1169. [Google Scholar] [CrossRef] [PubMed]
- Eacker, S.M.; Shima, J.E.; Connolly, C.M.; Sharma, M.; Holdcraft, R.W.; Griswold, M.D.; Braun, R.E. Transcriptional profiling of androgen receptor (AR) mutants suggests instructive and permissive roles of AR signaling in germ cell development. Mol. Endocrinol. (Baltimore, Md.) 2007, 21, 895–907. [Google Scholar] [CrossRef] [PubMed]
- De Gendt, K.; Swinnen, J.V.; Saunders, P.T.; Schoonjans, L.; Dewerchin, M.; Devos, A.; Tan, K.; Atanassova, N.; Claessens, F.; Lecureuil, C.; et al. A Sertoli cell-selective knockout of the androgen receptor causes spermatogenic arrest in meiosis. Proc. Natl. Acad. Sci. USA 2004, 101, 1327–1332. [Google Scholar] [CrossRef] [PubMed]
- Di Lorenzo, A.; Yang, Y.; Macaluso, M.; Bedford, M.T. A gain-of-function mouse model identifies PRMT6 as a NF-κB coactivator. Nucleic Acids Res. 2014, 42, 8297–8309. [Google Scholar] [CrossRef] [PubMed]
- Yoshimatsu, M.; Toyokawa, G.; Hayami, S.; Unoki, M.; Tsunoda, T.; Field, H.I.; Kelly, J.D.; Neal, D.E.; Maehara, Y.; Ponder, B.A.; et al. Dysregulation of PRMT1 and PRMT6, type I arginine methyltransferases, is involved in various types of human cancers. Int. J. Cancer J. Int. Cancer 2011, 128, 562–573. [Google Scholar] [CrossRef] [PubMed]
- Vieira, F.Q.; Costa-Pinheiro, P.; Ramalho-Carvalho, J.; Pereira, A.; Menezes, F.D.; Antunes, L.; Carneiro, I.; Oliveira, J.; Henrique, R.; Jeronimo, C. Deregulated expression of selected histone methylases and demethylases in prostate carcinoma. Endocr. Relat. Cancer 2014, 21, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Stein, C.; Riedl, S.; Ruthnick, D.; Notzold, R.R.; Bauer, U.M. The arginine methyltransferase PRMT6 regulates cell proliferation and senescence through transcriptional repression of tumor suppressor genes. Nucleic Acids Res. 2012, 40, 9522–9533. [Google Scholar] [CrossRef] [PubMed]
- Kang, N.; Chen, P.; Chen, Y.; Zeng, H.; He, X.; Zhu, Y. PRMT6 mediates CSE induced inflammation and apoptosis. Int. Immunopharmacol. 2015, 24, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Majumder, S.; Liu, Y.; Ford, O.H., 3rd; Mohler, J.L.; Whang, Y.E. Involvement of arginine methyltransferase CARM1 in androgen receptor function and prostate cancer cell viability. Prostate 2006, 66, 1292–1301. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.C.; Lin, S.R.; Luo, M.L.; Guo, H.; Wu, H.W.; Jiang, Z.M.; Gui, Y.T. Expression of a testis-specific gene 1700001022Rik in mice and its bioinformatic analysis. Natl. J. Androl. 2015, 21, 391–395. [Google Scholar]
- Yang, L.; Wang, Y.; Zhang, Q.; Lai, Y.; Li, C.; Huang, W.; Duan, Y.; Jiang, Z.; Li, X.; Cai, Z.; et al. Identification of HSF1 as a novel androgen receptor-regulated gene in mouse Sertoli cells. Mol. Reprod. Dev. 2014, 81, 514–523. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.H.; Ma, H.; Tan, T.Z.; Ng, S.S.; Soong, R.; Mori, S.; Fu, X.Y.; Zernicka-Goetz, M.; Wu, Q. Protein arginine methyltransferase 6 regulates embryonic stem cell identity. Stem Cells Dev. 2012, 21, 2613–2622. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, M.; Li, Y.; Guo, H.; Lin, S.; Chen, J.; Ma, Q.; Gu, Y.; Jiang, Z.; Gui, Y. Protein Arginine Methyltransferase 6 Involved in Germ Cell Viability during Spermatogenesis and Down-Regulated by the Androgen Receptor. Int. J. Mol. Sci. 2015, 16, 29467-29481. https://doi.org/10.3390/ijms161226186
Luo M, Li Y, Guo H, Lin S, Chen J, Ma Q, Gu Y, Jiang Z, Gui Y. Protein Arginine Methyltransferase 6 Involved in Germ Cell Viability during Spermatogenesis and Down-Regulated by the Androgen Receptor. International Journal of Molecular Sciences. 2015; 16(12):29467-29481. https://doi.org/10.3390/ijms161226186
Chicago/Turabian StyleLuo, Manling, Yuchi Li, Huan Guo, Shouren Lin, Jianbo Chen, Qian Ma, Yanli Gu, Zhimao Jiang, and Yaoting Gui. 2015. "Protein Arginine Methyltransferase 6 Involved in Germ Cell Viability during Spermatogenesis and Down-Regulated by the Androgen Receptor" International Journal of Molecular Sciences 16, no. 12: 29467-29481. https://doi.org/10.3390/ijms161226186