





Abstract
Objectives: To study the phenotype and phylogenetic relatedness of invasive Streptococcus pneumoniae strains isolated in Portugal.
Methods: A total of 614 invasive S. pneumoniae strains, isolated in Portugal from 1999 to the first 6 months of 2002, were characterized using serotyping and antimicrobial susceptibility testing. National outpatient sale data for macrolides were compared with erythromycin resistance. We investigated the main clonal lineages from erythromycin-resistant strains of serotype 14 by pulsed-field gel electrophoresis (PFGE) and multilocus sequence typing (MLST).
Results: The emergence of erythromycin-resistant strains correlated well with the usage of azithromycin in Portugal during the period of the study (r=0.900, P=0.001). Serotype 14 made the largest contribution to this emergence. We found two clonal complexes (CC) among erythromycin-resistant strains: CC1 was formed by three sequence types (ST), ST156, ST143 and ST1042, and CC2 by ST15 and ST9. All STs described, except ST1042, are putative founders of clonal groups or subgroups from serotype 14 as defined in the pneumococcal MLST database.
Conclusions: This study suggests that macrolides usage is an important factor enhancing the spread of invasive erythromycin-resistant S. pneumoniae clones in Portugal. The study also makes a contribution to the understanding of spread of erythromycin-resistant clones in an international context.
Introduction
The emergence of drug-resistant strains of Streptococcus pneumoniae is a growing problem worldwide.1 In Europe, the prevalence of macrolide resistance is increasing in non-invasive strains mainly in southern countries.2 In Portugal, the prevalence of erythromycin resistance of S. pneumoniae was stable during 1989–1993.3 To our knowledge, no further data have been published since. Molecular typing analyses show that only a small number of dominant clones are responsible for the spread of antibiotic resistance worldwide.4
We analysed macrolide usage and the genetic background of invasive S. pneumoniae clones of serotype 14 responsible for the increase in erythromycin resistance in Portugal. We also evaluated their relationship with drug-resistant clones in other countries.
Materials and methods
Clinical isolates
During January 1999–June 2002, 614 consecutive and non-repetitive invasive strains of S. pneumoniae were collected in the Antibiotic Resistance Unit at the National Institute of Health in Lisbon, in a multicentre study with 24 participating hospitals. The strains were isolated from blood, cerebrospinal fluid and pleural liquid from clinically ill patients.
Phenotypic characterization
For all isolates, MICs of penicillin (Wyeth Lederle Portugal, Algés, Portugal), cefotaxime and ceftriaxone (Roche Pharmaceuticals, Amadora, Portugal), tetracycline and chloramphenicol (Atral, Carregado, Portugal), erythromycin (Abbot Laboratórios, Amadora, Portugal), clindamycin (Laboratórios Upjohn Internacional, Lisboa, Portugal), ofloxacin (Farmoz, Prior Velho, Portugal) and ciprofloxacin (Bayer Portugal, Carnaxide, Portugal) were determined by an agar dilution method. MICs were interpreted according to NCCLS,5 except for ciprofloxacin (resistance was defined as a MIC value ≥4 mg/L). The MLSB phenotype was scored when simultaneous resistance to erythromycin and clindamycin was observed and the M phenotype when resistance to erythromycin only was observed. The M phenotype was confirmed by the induction test described previously.6 Serotypes were performed by Dot-blot and Quellung reaction using type-specific antiserum (Statens Serum Institut, Copenhagen, Denmark).7
Antimicrobial use data
National outpatient sale data [taken from International Medical Statistics (IMS Health, Portela, Portugal)] were used for macrolides/lincosamides (J01F) (azithromycin, clarithromycin, clindamycin, erythromycin, lincomycin, midecamycin, roxithromycin, spiramycin, telithromycin) for the period 1994–2002. The data were normalized to the Anatomic Therapeutic Classification system proposed by WHO.8 The number of packages consumed annually per 1000 inhabitants was used to enable the comparison of macrolide use over time in the same geographic area. The correlation between antimicrobial consumption and rates of macrolide resistance was assessed by the Spearman correlation coefficient applying the natural logarithm to the antimicrobial resistance, in order to have a linear relation. The null hypothesis was rejected for P values of <0.01.
Genotypic characterization
All erythromycin-resistant strains of serotype 14 (n=25) were investigated by PFGE and multilocus sequence typing (MLST) (eBURST).9,10 PFGE DNA fingerprints and MLST data were combined and analysed using Bionumerics software (version 3.0) (Applied Maths, St-Martens-Latem, Belgium). eBURST is an algorithm for displaying the relationships between closely related isolates of a bacterial species or population. This analysis was used to determine the main clonal complexes (CC) and the order of sequence types (ST) within CCs from MLST data.10 The algorithm predicts the descent from the founding genotype to the other genotypes in the group, displaying the output as a radial diagram, centred on the predicted founding genotypes. All strains of serotype 14 (n=274) in the pneumococcal MLST database were included in the analysis,10 to establish phylogenetic relationships between the erythromycin-resistant strains of this study and the principal CCs of serotype 14 already described.
Results and discussion
Phenotypic characterization
The MICs of 614 strains isolated in Portugal in 1999, 2000, 2001 and the first 6 months of 2002 were determined. The MICs of erythromycin were in the range 0.5–≥64 mg/L and for clindamycin in the range 2–≥64 mg/L. Overall, 13.6% and 10.1% of strains were resistant to erythromycin (MIC ≥0.5 mg/L) and clindamycin (MIC ≥0.5 mg/L), respectively. We showed an emerging macrolide resistance compared with the period 1989–1993:3 when 1.9% and 1.4% of strains were resistant to erythromycin and clindamycin, respectively. This observation is consistent with trends in other countries.2 In our study, the majority of erythromycin-resistant strains (74%) were also resistant to clindamycin and so had the MLSB phenotype, as in the period 1989–1993 (76%).
Most erythromycin-resistant strains were in one of three serotypes: 14 (31.0%), 6B (26.2%) and 19F (14.3%). The most common multidrug resistance phenotypes were: (i) penicillin, tetracycline, erythromycin and clindamycin, mainly for serotypes 6B, 19F, 14 (in descending order); (ii) tetracycline, erythromycin, clindamycin and chloramphenicol, for serotypes 6B and 19F; and (iii) penicillin, ceftriaxone, cefotaxime, tetracycline, erythromycin and clindamycin only for serotype 14.
Antimicrobial consumption and resistance
Outpatient sales of macrolides varied between 149.2 packages/1000 inhabitants/year in 1994 to 220.4 in 2002 (Figure 1a). Clarithromycin use was increased from 21.3 packages/1000 inhabitants/year in 1994 to 75.2 in 1999, and decreased until 2002 (58.8 packages/1000 inhabitants/year) (Figure 1a). Azithromycin has only been sold in Portugal since 1995 (0.85 packages/1000 inhabitants/year). Azithromycin use had a significant increase from 32.2 packages/1000 inhabitants/year in 1996 to 107.5 in 2002 (Figure 1a).
A strong correlation was found between the resistance to erythromycin and the use of azithromycin (r=0.900, P=0.001) (Figure 1b). These results suggest that the increase in azithromycin use is associated to the increase in macrolide resistance in S. pneumoniae.
Genotypic characterization
Genotypic characterization of the largest serotype contributor (serotype 14) showed that the increase in erythromycin resistance is due to clones included in two CCs, formed by five STs (Figures 2 and 3). The CC1 was formed by ST156, ST143 and ST1042 (Figure 2). MLST showed that strains of ST143 are double loci variants from ST156 (Figure 3a). Isolate 3569 is a previously undescribed single locus variant (SLV) of ST156, and was assigned to ST1042 (Figure 3a). CC2 consisted of ST15 and ST9 (Figure 2). ST9 is an SLV from ST15 (Figure 3b). eBURST analysis showed that CC1 and CC2 found in this study (Figure 2) are part of the lineages 3 (Figure 3a) and 2 (Figure 3b), respectively. These lineages were found among 121 STs (grouped into 10 lineages) from all serotype 14 strains in the pneumococcal MLST database.
Erythromycin-resistant clone ST156 present in Portugal is a serotype 14 variant of the erythromycin-susceptible international clone Spain9V−3 (Spain9V−3–14).9,11 This variant spread internationally, but most isolates are susceptible to macrolides, as identified in the MLST database (Figure 4).10 ST156 found in this study has been described in different countries, but in Uruguay it was described as being resistant to erythromycin.10,11 eBURST analysis of all strains of serotype 14 in the pneumococcal MLST database may indicate that ST156 was the first in the evolutionary pathway of CC1 (the ancestral type) (Figure 3a). The greater variability of PFGE and resistance patterns of ST156 than of ST143 in our sample is consistent with this possibility. ST144, not present among our strains (SLV variant of ST156), is a putative ancestor of ST143.
ST143 is the biggest clonal lineage in our sample (eight of 25 strains). This lineage has previously been described in Poland: all but one were erythromycin-susceptible isolates.10 Possibly this Portuguese clone was imported from that country. ST330 is an erythromycin-resistant clonal lineage, and is an SLV of ST143 (Figure 3a).10 eBURST analysis suggests that this ST, originally from a subpopulation of ST143 erythromycin-resistant strains, is probably of the same population as found in our study, and is genetically more distant from ST156. The new multidrug-resistant clonal lineage ST1042 could be a result of a recombinational event at the ddl locus.
Our ST9 strains had the same allelic profile and resistance pattern as England14−9, an international erythromycin-resistant clone.4 This clone of serotype 14 has been described in the UK, Italy, Argentina, Germany10 and in carriers in Portugal.12 In Australia, this clone was found in susceptible and resistant strains, as identified in the updated pneumococcal MLST database.10 The clonal lineage ST15 we found among Portuguese invasive strains has also been described in the UK, Taiwan, Italy, Hong Kong, Germany10 and in Portuguese carriers.12 Both susceptible and resistant strains of this ST have been described in Brazil, and susceptible strains have been found in the Netherlands.10 eBURST analysis shows that ST15 might be the ancestral type for CC2 (Figure 3b). This is consistent with the diversity of ST15 resistance patterns in our study (erythromycin, erythromycin plus clindamycin, penicillin plus erythromycin plus clindamycin, and tetracycline plus erythromycin plus clindamycin) compared with ST9 (only erythromycin resistance pattern). Possibly ST9 diverged from ST15 very recently, differing as attested by a single nucleotide polymorphism in the xpt gene sequence. This is reinforced with the PFGE data, which showed that ST15 and ST9 are phylogenetically very close.
Four STs described in this work (ST156, ST143, ST15 and ST9) are putative founders of major clonal groups and subgroups within serotype 14; they probably already existed in Portugal prior to the increase in erythromycin resistance. Both erythromycin-resistant and susceptible strains from invasive diseases are found in the same clonal lineage in several countries, including Portugal (data not shown). As the STs we describe include members of the major internationally disseminated clones of serotype 14 susceptible to macrolides, this also suggests that the resistance mechanisms were acquired locally (Figure 4).
To our knowledge, this study is the first report of the genetic relatedness of invasive pneumococcal clones responsible for the emergence of erythromycin resistance in Portugal. This study suggests that azithromycin use is an important selection factor for the spread of these clones in Portugal, which is also associated with the increase in macrolide resistance. This also provides a contribution to the understanding of the spread of these antibiotic-resistant clones in an international context.
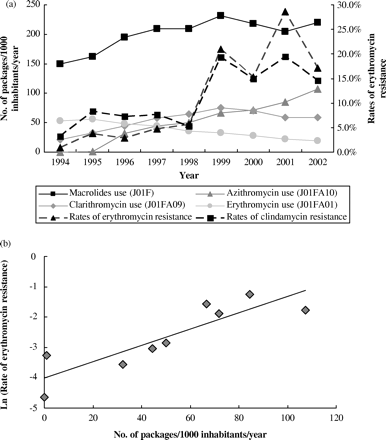
(a) Antimicrobial usage and resistance. (b) Correlation between azithromycin use and rates of erythromycin resistance, during 1994–2002, using Spearman's correlation coefficient (r=0.900, P=0.001). Source: rates of antimicrobial resistance (M. Caniça).
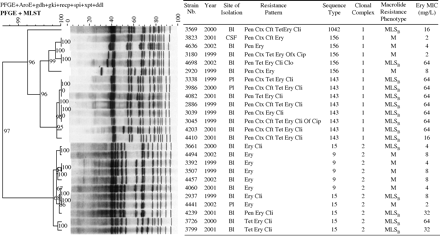
Properties and phylogenetic relationships based on MLST and PFGE data from 25 erythromycin-resistant strains with serotype 14. The Dice band-based similarity coefficient was used for PFGE pattern analysis, with a band position tolerance of 1% and an optimization of 1%. PFGE DNA fingerprints and MLST data were combined and analysed using Bionumerics software. The dendrogram was constructed by the unweighted pair-group method with average linkages using the composite data set of MLST and the PFGE patterns. Bl, blood; Pl, pleural liquid; CSF, cerebrospinal fluid; Pen, penicillin; Ctx, cefotaxime; Cft, ceftriaxone; Tet, tetracycline; Ery, erythromycin; Cli, clindamycin; Clo, chloramphenicol; Of, ofloxacin; Cip, ciprofloxacin.
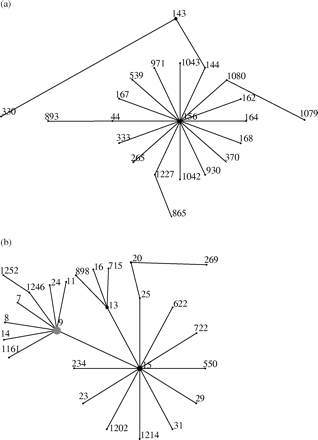
Relationship between 25 erythromycin-resistant strains of serotype 14 from this study and serotype 14 strains from the pneumococcal MLST database by eBURST analysis. (a) Lineage 3 from eBURST analysis: ST156 is the ancestral type. Clonal complex 1 (ST156, ST143 and ST1042, from this study) is included in this lineage. (b) Lineage 2 from eBURST analysis: ST15 is the ancestral type. Clonal complex 2 (ST15 and ST9, from this study) is included in this lineage; the black circle in the centre of the eBURST diagrams predicts primary founders; the grey circle defines a subgroup founder; Numbers on the diagram correspond to the STs.
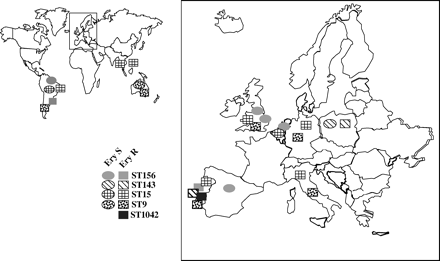
International distribution of pneumococcal clones of serotype 14 found in Portugal. Also shown is a local clone, ST1042. Ery S, erythromycin-susceptible clones; Ery R, erythromycin-resistant clones. ST, sequence type. Source: Pneumococcal MLST database.10
We wish to thank Deolinda Louro for expert technical assistance with MIC determinations, and the hospitals participating in GEMVSA (A Portuguese Multicenter Study Group) for providing strains of S. pneumoniae. We acknowledge IMS Health, in Portugal, for data of antimicrobial consumption and Baltazar Nunes (National Observatory for Health, NIH, Portugal) for his help in statistical analysis. We also acknowledge use of the pneumococcal MLST database, which is located at Imperial College London and is funded by the Wellcome Trust. This work was supported in part by Comissão de Fomento da Investigação em Cuidados de Saúde grant 237/99 from Ministério da Saúde, Portugal. Serotyping study was supported by a Wyeth Lederle Portugal (Farma) grant. Ricardo Dias was supported by the Portuguese ‘NIH Scientific Research Ricardo-Jorge’ grant. This work was presented in part at the Sixth International Meeting on Microbial Epidemiological Markers, Les Diablerets, Switzerland, 2003, and at the Fourth International Symposium on Pneumococci and Pneumococcal Diseases, Helsinki, Finland, 2004.
References
Appelbaum, P. (
Jacobs, M., Felmingham, D., Appelbaum, P. et al. (
Pato, M., Carvalho, C. & Tomasz, A. (
McGee, L., McDougal, L., Zhou, J. et al. (
National Committee for Clinical Laboratory Standards. (
Seppälä, H., Nissinen, A., Yu, Q. et al. (
Fenoll, A., Jado, I., Vicioso, D. et al. (
W.H.O. Collaborating Centre for Drug Statistics Methodology. (
Caniça, M., Dias, R., Vaz-Pato, M. et al. (
Pneumococcal MLST database. [Online.] http://www.mlst.net/ (24 May 2004, date last accessed).
Coffey, T., Daniels, M., Enright, M. et al. (
Author notes
Antibiotic Resistance Unit, Centre of Bacteriology, National Institute of Health Dr. Ricardo Jorge, Av. Padre Cruz, 1649–016 Lisboa, Portugal

{kind=link}
{kind=link}
{kind=link}
{kind=link}