




Abstract
BACKGROUND: Endometriosis is a gynaecological disease that can be associated with severe pelvic pain; however, the mechanisms by which pain is generated remain unknown. METHODS: Peritoneal endometriotic lesions and normal peritoneum were prepared from women with and without endometriosis (n = 40 and 36, respectively). Specimens were also prepared from endosalpingiosis lesions (n = 9). These sections were stained immunohistochemically with antibodies against protein gene product 9.5, neurofilament (NF), nerve growth factor (NGF), NGF receptor p75 (NGFRp75), substance P (SP), calcitonin gene-related peptide (CGRP), acetylcholine (ACh) and tyrosine hydroxylase (TH) to demonstrate myelinated, unmyelinated, sensory, cholinergic and adrenergic nerve fibres. RESULTS: There were significantly more nerve fibres identified in peritoneal endometriotic lesions than in normal peritoneum (P < 0.001) or endosalpingiosis lesions (P < 0.001). These nerve fibres were SP, CGRP, ACh or TH immunoreactive. Many of these markers were co-localized. There was an intense NGF immunoreactivity near endometriotic glands, and NGFRp75 immunoreactive nerve fibres were present near endometriotic glands and blood vessels in the peritoneal endometriotic lesions. CONCLUSIONS: Peritoneal endometriotic lesions were innervated by sensory Aδ, sensory C, cholinergic and adrenergic nerve fibres. These nerve fibres may play an important role in the mechanisms of pain generation in this condition.
Introduction
Endometriosis is a gynaecological disease defined by the histologic presence of endometrial glands and stroma outside the uterine cavity, most commonly implanted over visceral and peritoneal surfaces within the female pelvis. This disease is a benign disorder; however, it exhibits cellular proliferation, cellular invasion and neoangiogenesis (Giudice et al., 1998). Although the exact prevalence of endometriosis in the general population is not clear, the prevalence in women of reproductive age is estimated to range between 10 and 15% (Lebovic et al., 2001). Endometriosis occurs throughout the pelvic cavity including pelvic organs such as the ovaries, pelvic peritoneum, uterus, Fallopian tube, vagina, rectovaginal septum, intestinal tract and the ureter.
Endometriosis is usually associated with pelvic pain such as chronic dysmenorrhoea, intermenstrual abdominal and pelvic pain, back pain, dysuria, dyschezia and dyspareunia. However, the relationship between pain and endometriosis is not well understood.
Several researchers have investigated the presence of nerve fibres in endometriotic lesions. Tamburro et al. (2003) demonstrated nerve fibres that expressed transforming growth factor-β1 (TGF-β1) in endometriotic lesions from women with dysmenorrhoea associated with endometriosis. Tulandi et al. (2001) demonstrated neurofilament (NF) immunoreactive nerve fibres in endometriotic lesions in women with endometriosis, and they reported that the distance between endometrial glands and nerve fibres in endometriotic lesions from women with pain was closer than in women with no pain. Berkley et al. (2004) demonstrated nerve fibres stained with calcitonin gene-related peptide (CGRP), substance P (SP) and vesicular monoamine transporter in endometriotic lesions in a rat model. We have recently demonstrated the unexpected finding of multiple small unmyelinated nerve fibres in the eutopic endometrium of women with laparoscopically confirmed endometriosis but not in the endometrium of women without endometriosis (Tokushige et al., 2006).
We have studied innervation with different types of specific immunohistochemical neuronal markers in human peritoneal endometriotic lesions from women with visually and biopsy proven endometriosis. Markers included polyclonal rabbit anti-protein gene product 9.5 (PGP9.5) (Lundberg et al., 1988), monoclonal mouse anti-human NF (Schlaepfer, 1987), polyclonal rabbit anti-SP (Berkley et al., 2004), rabbit anti-CGRP (Berkley et al., 2004), polyclonal rabbit anti-acetylcholine (anti-ACh) (Janossy et al., 1998) and monoclonal anti-tyrosine hydroxylase (anti-TH) (Calder et al., 1998) for myelinated, unmyelinated, sensory Aδ, sensory C, cholinergic and adrenergic nerve fibres. PGP9.5 is a highly specific pan-neuronal marker and is also expressed in neuroendocrine cells. NF is a highly specific marker for myelinated nerve fibres. SP and CGRP are sensory nerve fibre markers, and they can be present in both Aδ and C nerve fibres. ACh is present in sympathetic preganglionic neurons and parasympathetic preganglionic and postganglionic neurons, and it is a specific marker for cholinergic nerve fibres. TH is present in sympathetic postganglionic neurons, and it is a specific marker for adrenergic nerve fibres. There is no cross-reactivity in these antibodies. We also used polyclonal rabbit anti-nerve growth factor (anti-NGF) and monoclonal mouse anti-human NGF receptor p75 (NGFRp75), a low-affinity NGFR, to find out whether these substances were associated with the development and survival of sensory, cholinergic and adrenergic nerve fibres in peritoneal endometriotic lesions in women with endometriosis. To demonstrate the relationship between nerve fibres and blood vessels, we performed double staining using PGP9.5 and CD34, a marker for endothelial cells.
We also studied nerve fibres in peritoneal lesions of endosalpingiosis to see whether this related and commonly co-existent pathological process increased nerve density in peritoneal endometriotic lesions. Endosalpingiosis is defined as the presence of epithelial inclusions, typified by tubal-like epithelium, with no endometrial cytogenic stroma, and the common presence of microcalcification (psammoma bodies). Such lesions most commonly involve the peritoneal surface of the uterus, Fallopian tubes, ovaries or Pouch of Douglas.
Materials and methods
Collection of tissue
This study was approved by the Human Ethics Committees of the Central Sydney Area Health Service and the University of Sydney, and all women gave their informed consent for participation.
Peritoneal endometriosis tissue was collected from 40 women with endometriosis (mean age 33.2 years; range 16–46 years) and normal peritoneum was also collected from 36 women without endometriosis (mean age 34.9 years; range 17–57 years) who underwent laparoscopy combined with hysteroscopy. The endometriosis patients all complained of dysmenorrhoea and a range of related pain symptoms. The 36 normal peritoneum tissue samples collected from women without laparoscopic evidence of endometriosis were from patients undergoing tubal sterilization, assessment before tubal reanastomosis or investigation of infertility (without any recognizable pathology).
Peritoneal endosalpingiosis lesions were identified in biopsy material from nine women (mean age 27.6 years; range 14–44 years) who had undergone laparoscopy combined with hysteroscopy for proven or suspected endometriosis.
The presence and nature of the endometriosis and endosalpingiosis lesions in biopsies were confirmed by an experienced gynaecological pathologist (P.R.).
The severity of pain was not assessed systematically and prospectively in this preliminary observational study, but detailed clinical information was recorded in a standard format. All endometriosis patients were staged according to the revised American Fertility Society (AFS) score (American Society for Reproductive Medicine, 1996), and ranged from stages I to IV. None of the women had received medical therapy for endometriosis before endoscopy and tissue sampling.
Immunohistochemistry
After resection, the specimens were immediately fixed in 10% neutral buffered formalin for ∼18–24 h, processed and embedded in paraffin wax according to a standard protocol. Each section was cut at 4 µm and routinely stained with haematoxylin and eosin. Antigen retrieval techniques for PGP9.5, NF and NGFRp75 were used. Serial sections, cut at 4 µm, were immunostained using antibodies for polyclonal rabbit anti–PGP9.5 (dilution 1 : 700; Dako Australia, Sydney, Australia) (a highly specific pan-neuronal marker) (Lundberg et al., 1988), monoclonal mouse anti-human NF protein (dilution 1 : 200; Dako Australia) (a highly specific marker for myelinated nerve fibres) (Schlaepfer, 1987), polyclonal rabbit anti-human NGF (dilution 1 : 1200; Santa Cruz Biotechnology, CA, USA), monoclonal mouse anti-human NGFRp75 (dilution 1 : 700; Dako Australia) (a low-affinity NGFR) and polyclonal rabbit anti-ACh (dilution 1 : 2500; Chemicon, CA, USA) for 30 min at room temperature. Serial sections were also immunostained with rabbit anti-SP (dilution 1 : 4000; Serotec, NC, USA), polyclonal rabbit anti-CGRP (dilution 1 : 20 000; Sigma, MO, USA) and TH (dilution 1 : 1200; Sigma) overnight at 4°C. Sections stained with PGP9.5, NF, NGFRp75, TH and ACh were washed in Tris Buffer, incubated with Envision-labelled polymer-AP mouse/rabbit (Dako Australia) for 30 min and stained with permanent fast red chromogen (Dako Australia) for 10 min. Sections stained with NGF, SP and CGRP were washed in Tris Buffer, incubated with Envision+ dual link system peroxidase (Dako Australia) for 30 min and stained with Liquid 3,3′-diaminobenzidine (DAB) + Substrate (Dako Australia) for 10 min.
For double staining, serial sections were immunostained with CD34 (dilution 1 : 700; Dako Australia) for 30 min at room temperature, washed in Tris Buffer, incubated with Envision+ dual link system peroxidase for 30 min and stained with Liquid DAB + Substrate for 10 min. The sections were then treated with acid block (Dako Australia) for 3 min and stained with polyclonal rabbit anti-PGP9.5 (dilution 1 : 500) for 30 min, washed in Tris Buffer, incubated with Envision-labelled polymer-AP mouse/rabbit for 30 min and stained with permanent fast red chromogen for 10 min.
All immunostaining was carried out on a Dako Autostainer Model S3400 (Dako USA, Carpinteria, CA, USA). Images of the sections were captured using Olympus microscope BX51 and digital camera DP70 (Olympus, Tokyo, Japan). We used normal skin as a positive control as it reliably contains myelinated and unmyelinated nerve fibres and endothelial cells expressing PGP9.5, NF, NGF, NGFRp75, SP, CGRP, ACh, TH and CD34. The functional layer of eutopic endometrium from women without endometriosis was used as a negative control as it does not contain any nerve fibres expressing PGP9.5, NF, NGF, NGFRp75, SP, CGRP, ACh and TH.
Statistical analysis
The images were captured using Olympus microscope BX51 and digital camera DP70, and an assessment of nerve fibre density was performed by Image Pro Plus Discovery (MediaCybernetics, MD, USA). Once the images’ features were acquired, an orthogonal grid mask was sketched above the original images. The sections of the grid were 250 µm per side. Once the grid was in position, nerve fibres in the peritoneal endometriotic lesions, peritoneal endosalpingiosis lesions and normal peritoneum, and the total number of squares covering the sections of peritoneal endometriotic lesions, peritoneal endosalpingiosis lesions and normal peritoneum were counted. The total number of nerve fibres was divided by the total number of squares covering the peritoneal endometriotic lesions, peritoneal endosalpingiosis lesions or the normal peritoneum to obtain an average of nerve fibres per square (each square of 250 × 250 µm). The results were expressed as the mean (±SD) number of nerve fibres per mm2 in each specimen from all peritoneal endometriotic lesions, peritoneal endosalpingiosis lesions and normal peritoneum samples, stained with the polyclonal antibody against PGP9.5 and the monoclonal antibody against NF. The counting procedure was carried out twice by two independent observers (each blinded to the other) without any knowledge of the clinical parameters or other prognostic factors. The concordance rate was >95% between the observers. Nerve fibre density between peritoneal endometriotic lesions from endometriosis patients, peritoneal endosalpingiosis lesions from patients with endosalpingiosis alone or patients with both endometriosis and endosalpingiosis and normal peritoneum from women without endometriosis was compared using the Kruskal–Wallis test and the Mann–Whitney test. Differences were considered to be significant at P < 0.05.
Nerve fibres stained with SP, CGRP, TH, ACh, NGF and NGFRp75 were assessed semiquantitatively due to the partial cross-reactivity of some of these markers with some blood vessels and connective tissues. Their staining pattern was compared with that of PGP9.5 to obtain a semiquantitative representation of these nerve fibres. Grading of density of these nerve fibres was represented as the percentage of total nerve fibres demonstrated by PGP9.5 immunoreactivity. + few (10–20%), ++ moderate (20–70%), +++ many (70–90%), ++++ abundant (90–100%).
In one series of experiments, serial sections were sequentially stained with all the markers listed above to assess the extent of co-localization of the markers around small nerve tracts within peritoneal endometriotic lesions and normal peritoneum.
Results
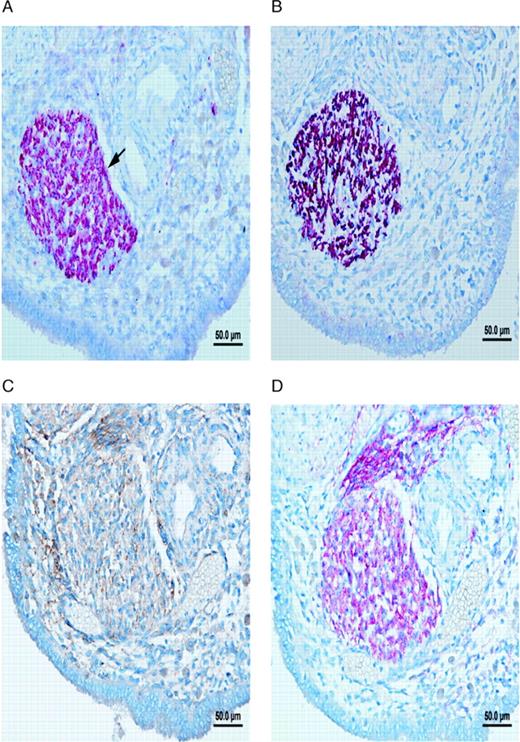
There were more nerve fibres stained with PGP9.5 in the peritoneal endometriotic lesions (n = 40) from women with endometriosis (mean 16.3 ± 10.0/mm2; range 6.8–53.9/mm2) than in normal peritoneum (n = 36) from women without endometriosis (mean 2.5 ± 1.3/mm2; range 0.5–5.5/mm2; P < 0.001; Table I). These nerve fibres were more frequently seen near endometriotic glands and blood vessels than elsewhere in the stroma (Figure 1A).
A quantitative and semiquantitative assessment of the density of different types of nerve fibres in peritoneal endometriotic lesions
| Normal peritoneum | Peritoneal endometriotic lesion | |
|---|---|---|
| Number of specimens | 36 | 40 |
| Total nerve fibre density (mean ± SD/mm2) (stained with PGP9.5) | 2.5 ± 1.3 | 16.3 ± 10.0 |
| Total nerve fibre density (mean ± SD/mm2) stained with NF | 1.0 ± 0.8 | 6.7 ± 3.7 |
| Presence of SP immunoreactive nerve fibres | ++ | ++ |
| Presence of CGRP immunoreactive nerve fibres | ++ | ++ |
| Presence of TH immunoreactive nerve fibres | ++ | ++ |
| Presence of ACh immunoreactive nerve fibres | + | + |
| Presence of NGFRp75 immunoreactive nerve fibres | ++++ | ++++ |
| Presence of NGF immunoreactive nerve fibres | ++++ | ++++ |
| Normal peritoneum | Peritoneal endometriotic lesion | |
|---|---|---|
| Number of specimens | 36 | 40 |
| Total nerve fibre density (mean ± SD/mm2) (stained with PGP9.5) | 2.5 ± 1.3 | 16.3 ± 10.0 |
| Total nerve fibre density (mean ± SD/mm2) stained with NF | 1.0 ± 0.8 | 6.7 ± 3.7 |
| Presence of SP immunoreactive nerve fibres | ++ | ++ |
| Presence of CGRP immunoreactive nerve fibres | ++ | ++ |
| Presence of TH immunoreactive nerve fibres | ++ | ++ |
| Presence of ACh immunoreactive nerve fibres | + | + |
| Presence of NGFRp75 immunoreactive nerve fibres | ++++ | ++++ |
| Presence of NGF immunoreactive nerve fibres | ++++ | ++++ |
PGP9.5, protein gene product 9.5; NF, neurofilament; SP, substance P; CGRP, calcitonin gene-related peptide; TH, tyrosine hydroxylase; ACh, acetylcholine; NGFRp75, nerve growth factor receptor p75; NGF, nerve growth factor.
Grading of density of these nerve fibres was represented as the percentage of total nerve fibres demonstrated by PGP9.5 immunoreactivity. +, few (10–20%); ++, moderate (20–70%); +++, many (70–90%); ++++, abundant (90–100%).
A quantitative and semiquantitative assessment of the density of different types of nerve fibres in peritoneal endometriotic lesions
| Normal peritoneum | Peritoneal endometriotic lesion | |
|---|---|---|
| Number of specimens | 36 | 40 |
| Total nerve fibre density (mean ± SD/mm2) (stained with PGP9.5) | 2.5 ± 1.3 | 16.3 ± 10.0 |
| Total nerve fibre density (mean ± SD/mm2) stained with NF | 1.0 ± 0.8 | 6.7 ± 3.7 |
| Presence of SP immunoreactive nerve fibres | ++ | ++ |
| Presence of CGRP immunoreactive nerve fibres | ++ | ++ |
| Presence of TH immunoreactive nerve fibres | ++ | ++ |
| Presence of ACh immunoreactive nerve fibres | + | + |
| Presence of NGFRp75 immunoreactive nerve fibres | ++++ | ++++ |
| Presence of NGF immunoreactive nerve fibres | ++++ | ++++ |
| Normal peritoneum | Peritoneal endometriotic lesion | |
|---|---|---|
| Number of specimens | 36 | 40 |
| Total nerve fibre density (mean ± SD/mm2) (stained with PGP9.5) | 2.5 ± 1.3 | 16.3 ± 10.0 |
| Total nerve fibre density (mean ± SD/mm2) stained with NF | 1.0 ± 0.8 | 6.7 ± 3.7 |
| Presence of SP immunoreactive nerve fibres | ++ | ++ |
| Presence of CGRP immunoreactive nerve fibres | ++ | ++ |
| Presence of TH immunoreactive nerve fibres | ++ | ++ |
| Presence of ACh immunoreactive nerve fibres | + | + |
| Presence of NGFRp75 immunoreactive nerve fibres | ++++ | ++++ |
| Presence of NGF immunoreactive nerve fibres | ++++ | ++++ |
PGP9.5, protein gene product 9.5; NF, neurofilament; SP, substance P; CGRP, calcitonin gene-related peptide; TH, tyrosine hydroxylase; ACh, acetylcholine; NGFRp75, nerve growth factor receptor p75; NGF, nerve growth factor.
Grading of density of these nerve fibres was represented as the percentage of total nerve fibres demonstrated by PGP9.5 immunoreactivity. +, few (10–20%); ++, moderate (20–70%); +++, many (70–90%); ++++, abundant (90–100%).
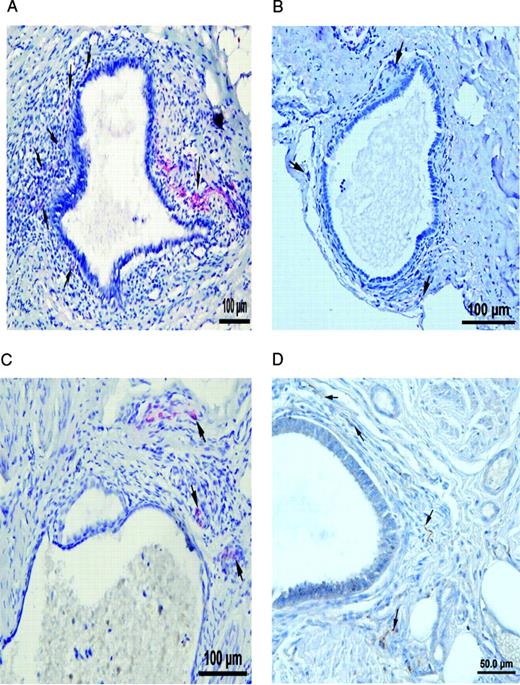
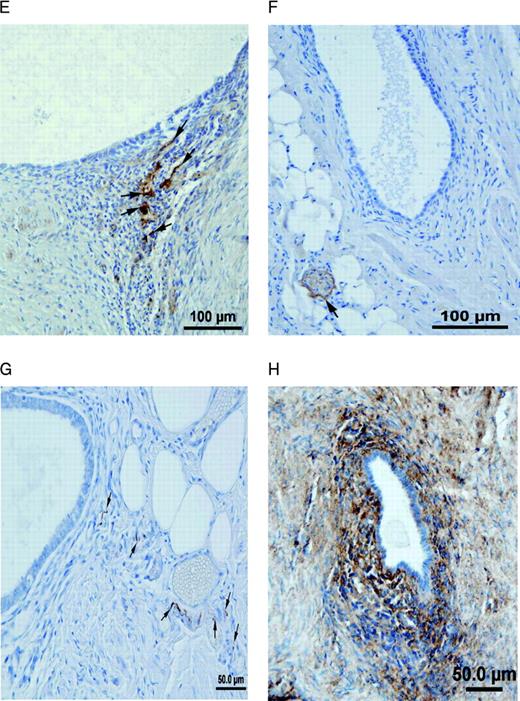
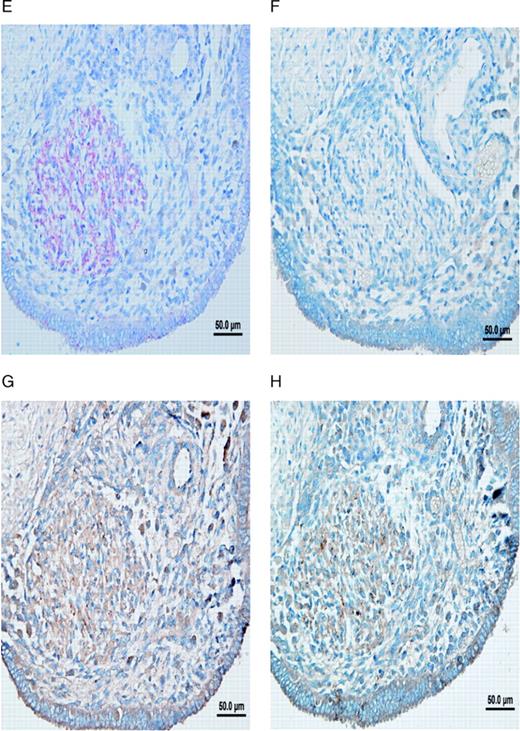
Nerve fibres in peritoneal endometriotic lesions and a peritoneal endosalpingiosis lesion. (A) Peritoneal endometriotic lesion stained with protein gene product 9.5 (PGP9.5) and permanent fast red chromogen (magnification ×200). Arrows denote PGP9.5-positive nerve fibres near an endometriotic gland. (B) Peritoneal endosalpingiosis lesion stained with PGP9.5 and permanent fast red chromogen (magnification ×200). Arrows denote tiny PGP9.5-positive nerve fibres. (C) Peritoneal endometriotic lesion stained with neurofilament (NF) and permanent fast red chromogen (magnification ×200). Arrows denote NF-positive myelinated nerve fibres near an endometriotic gland. (D) Peritoneal endometriotic lesion stained with substance P (SP) and 3,3′-diaminobenzidine (DAB) (magnification ×400). Arrows denote tiny SP-positive nerve fibres near an endometriotic gland. (E) Peritoneal endometriotic lesion stained with calcitonin gene-related peptide (CGRP) and DAB (magnification ×200). Arrows denote CGRP-positive nerve fibres near an endometriotic gland. (F) Peritoneal endometriotic lesion stained with acetylcholine (ACh) and DAB (magnification ×200). Arrow denotes ACh-positive nerve fibres near an endometriotic gland. (G) Peritoneal endometriotic lesion stained with tyrosine hydrolase (TH) and DAB (magnification ×400). Arrows denote TH-positive nerve fibres near an endometriotic gland. (H) Peritoneal endometriotic lesion stained with nerve growth factor (NGF) and DAB (magnification ×200). (I) Peritoneal endometriotic lesion stained with nerve growth factor receptor p75 (NGFRp75) and permanent fast red chromogen (magnification ×200). Black arrows denote NGFRp75-positive nerve fibres and nerve fibre trunks near an endometriotic gland (red arrow). Magnification ×400 was used due to small size of nerve fibres. Scale bars represent 50 µm in D, G and H; and 100 µm in A–C, E, F and I.
Nerve fibres were present in greater density in peritoneal endometriotic lesions (mean 16.3 ± 10.0/mm2; range 6.8–53.9/mm2) than in peritoneal endosalpingiosis lesions (n = 9) (mean 3.8 ± 0.9/mm2; range 2.7–5.8/mm2; Figure 1B; P < 0.001). The density of nerve fibres in endosalpingiosis lesions was also significantly greater than in normal peritoneum (P < 0.002).
There were also more NF immunoreactive nerve fibres in peritoneal endometriotic lesions (6.7 ± 3.7/mm2; range 2.0–20.0/mm2) than in normal peritoneum (1.0 ± 0.8/mm2; range 0–2.7/mm2) and peritoneal endosalpingiosis (1.8 ± 0.4/mm2; range 1.3–2.8/mm2; P < 0.001; Figure 1C; Table I). However, only 4 out of 40 (10%) peritoneal endometriotic lesions showed NF immunoreactive nerve fibres near to endometriotic glands.
With confirmation by the specific markers for SP, CGRP, ACh and TH, nerve fibres in the peritoneal endometriotic lesions were demonstrated to be sensory Aδ, sensory C, cholinergic and adrenergic nerve fibres (Figure 1D–G, Table I).
There was an intense NGF immunoreactivity near endometriotic glands in peritoneal endometriotic lesions (Figure 1H). More than 90% of the total PGP9.5 immunoreactive nerve fibres were stained with NGF and NGFRp75 in peritoneal endometriotic lesions (Figure 1I).
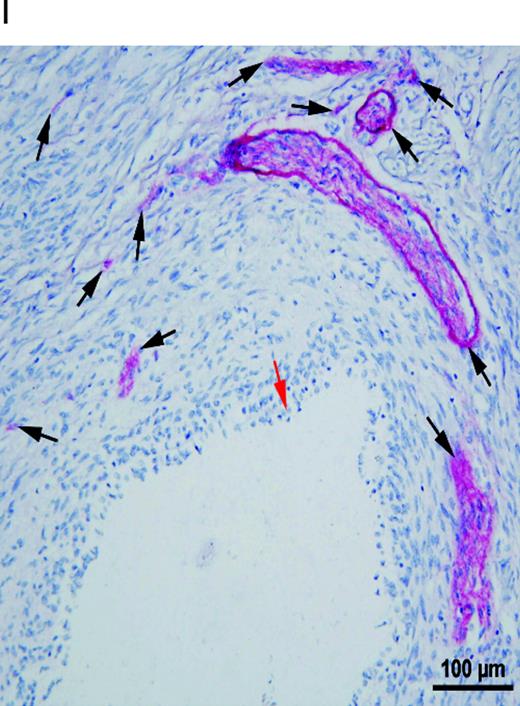
Serial sections stained with PGP9.5, NF, NGF, NGFRp75, TH, ACh, SP, CGRP and CD34 showed that several nerve fibres in peritoneal endometriotic lesions showed co-localization of SP, CGRP, TH, NGF and NGFRp75. Those nerve fibres were a mixture of sensory C, sensory Aδ and adrenergic nerve fibres (Figure 2A–H) and associated with blood vessels (Figure 2I). Nerve fibres stained with ACh were not co-localized with SP, CGRP, TH, NGF and NGFRp75.
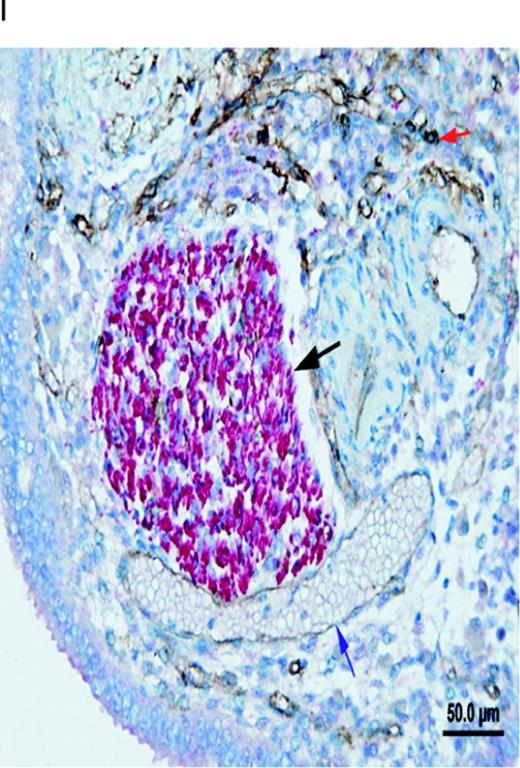
Different types of nerve fibres in a peritoneal endometriotic lesion (serial sections). (A) Peritoneal endometriotic lesion showing a large nerve trunk stained with protein gene product 9.5 (PGP9.5) (magnification ×400). Nerve fibres were stained red with permanent fast red chromogen. (B) Peritoneal endometriotic lesion from the same patient stained with neurofilament (NF) (magnification ×400) as nerve fibres were stained red with permanent fast red chromogen. (C) Peritoneal endometriotic lesion from the same patient stained with nerve growth factor (NGF) and 3,3′-diaminobenzidine (DAB) (magnification ×400). Nerve fibres and some surrounding tissues were weakly stained brown. (D) Peritoneal endometriotic lesion from the same patient stained with nerve growth factor receptor p75 (NGFRp75) and permanent fast red chromogen (magnification ×400). Nerve fibres and some closely localized and related surrounding tissues were stained red. (E) Peritoneal endometriotic lesion from the same patient stained with tyrosine hydrolase (TH) and permanent fast red chromogen (magnification ×400). Nerve fibres were weakly stained red. (F) Peritoneal endometriotic lesion from the same patient stained with acetylcholine (ACh) and DAB (magnification ×400). No nerve fibres were stained. (G) Peritoneal endometriotic lesion from the same patient stained with substance P (SP) and DAB (magnification ×400). Nerve fibres were weakly stained brown. (H) Peritoneal endometriotic lesion from the same patient stained with calcitonin gene-related peptide (CGRP) and DAB (magnification ×400). Nerve fibres were weakly stained brown. (I) Peritoneal endometriotic lesion from the same patient double-stained with PGP9.5 and CD34 (magnification ×400). The nerve trunk was intensely stained red (black arrow) and blood vessels (blue and red arrows) were stained brown. Red arrow denotes high density of capillaries and blue arrow denotes thin walled blood vessel. Scale bars represent 50 µm.
Discussion
This study has demonstrated multiple, small unmyelinated nerve fibres in peritoneal endometriotic lesions in 40 women with confirmed endometriosis. The density of nerve fibres in peritoneal endometriotic lesions was much greater than in normal peritoneum in women with no endometriosis and much greater than in lesions of endosalpingiosis. It was somewhat surprising that the density of nerve fibres in endosalpingiosis lesions was no different from that in normal peritoneum, suggesting that increased nerve fibre density in endometriosis lesions and in the eutopic endometrium of endometriosis patients (Tokushige et al., 2006) is a specific finding for that condition rather than a general response of peritoneum to development of a pathological lesion. Nerve fibres were more frequently seen near endometriotic glands in peritoneal endometriotic lesions. Only 4 out of 40 (10%) peritoneal endometriotic lesions showed NF immunoreactive (myelinated) nerve fibres near endometriotic glands, and NF immunoreactive nerve fibres demonstrated in the other 36 peritoneal endometriotic lesions were distant from endometriotic glands. Therefore, most of the nerve fibres demonstrated in close proximity to endometriotic glands were unmyelinated. Those nerve fibres stained with NF in four peritoneal endometriotic lesions may be Aδ fibres that transmit sharp, pricking localized pain to the central nervous system.
Confirmation by the specific markers for sensory, cholinergic and adrenergic nerve fibres demonstrated that single trunks and nerve fibre trunks in peritoneal endometriotic lesions were found to contain sensory Aδ, sensory C, cholinergic and adrenergic nerve fibres. Moderate numbers of sensory C and adrenergic nerve fibres were present near endometriotic glands. The other markers for sensory Aδ and cholinergic nerve fibres were not obviously located in close relation to glands. Several single trunks and nerve fibre trunks in peritoneal endometriotic lesions showed co-localization of SP, CGRP, TH and NF, and those nerve fibre trunks were a mixture of sensory C, sensory Aδ and adrenergic nerve fibres. Nerve fibres stained with ACh were not co-localized with SP, CGRP and TH.
Tulandi et al. (2001) demonstrated NF immunoreactive nerve fibres in peritoneal endometriotic lesions and reported that the distance between endometriotic glands and nerve fibres in endometriotic lesions from women with pain was closer than in women with no pain. We also demonstrated SP, CGRP, TH, NF and ACh immunoreactive nerve fibres close to endometriotic glands in women with pain associated with endometriosis.
It is not yet clear what neural stimuli may initiate the severe pain sensations that some women experience with endometriosis. Endometriosis is an inflammatory disease and many inflammatory mediators are released from damaged cells and tissues in the vicinity of endometriotic lesions. These substances include histamine, serotonin (5-HT), bradykinin, prostaglandins (PGs), leukotrienes, interleukin (IL)-1β, IL-6, IL-8, ACh, vascular endothelial growth factor (VEGF), tumour necrosis factor-α (TNF-α), endothelins, epidermal growth factor (EGF), TGF-β, platelet-derived growth factor (PDGF) and NGF (Julius et al., 2001; Mantyh, 2002; Richardson and Vasko, 2002). Molecules secreted directly from endometriotic lesions include PGs (Ylikorkala and Viinikka, 1983; De Leon et al., 1988), TNF-α (Bergqvist et al., 2001) and NGF (Anaf et al., 2002). These substances can all sensitize and/or activate sensory nerve endings by interacting with cell-surface receptors (Mantyh, 2002; Schaible et al., 2002). It is clear that many peritoneal endometriotic lesions are locally tender and this may be a manifestation of an inflammatory reaction. Palter et al. (1996) first described active endometriotic lesions as a source of pain using ‘conscious pain mapping’. Almeida and Val-Gallas (1997) found 48 positive findings in 50 women (96%) with conscious pain mapping, and Demco (1998) also found that most women localized or mapped their pain to their endometriotic lesions.
Tamburro et al. (2003) demonstrated TGF-α1 immunoreactive nerve fibres in peritoneal endometriotic lesions from women with dysmenorrhoea. Because TGF-α1 can increase cyclooxygenase-2 activity (Fong et al., 2000), TGF-α1 may increase PG production in sensory and sympathetic nerve fibres in women with endometriosis, constituting one further possible mechanism of pain generation.
When sensory nerve fibres are stimulated by inflammatory substances listed above, neurotransmitters such as SP, CGRP, endothelin, histamine, glutamate, PGs and vaso-intestinal peptide can be secreted from sensory nerve endings (Mantyh, 2002; Liddle and Nathan, 2004). SP and CGRP can contribute to the inflammatory response by causing vasodilation, plasma extravasation (leakage of proteins and fluid from postcapillary venules) and cellular infiltration by interacting with endothelial cells, arterioles, mast cells, neutrophils and immune cells (Maggi, 1995). SP can also act on mast cells in the vicinity of sensory nerve endings to evoke degranulation and the release of TNF-α, histamine, PGD2, IL-8 and leukotriene B4, as well as acting on platelets to release 5-HT, providing a positive feedback (Steinhoff et al., 2003). CGRP has a wide range of biological activities, including sensory transmission (Gibson et al., 1984), regulation of glandular secretion (Onuoha and Alpar, 2001), proliferative effect on human endothelial cells (Onuoha and Alpar, 2001) and inhibiting SP degradation by a specific endopeptidase (Le Greves et al., 1985) to enhance SP release, thereby amplifying the effects (Steinhoff et al., 2003).
C nerve fibres can develop a greater sensitivity to norepinephrine (NE) during inflammation (Banik et al., 2001). Sympathetic postganglionic neurons (adrenergic nerve terminals) can synthesize PGE2 and PGI2 (Gonzales et al., 1989). Taiwo et al. (1990) reported that the hyperalgesia induced by NE is mediated by PGs produced by sympathetic postganglionic neuron terminals, as NE is considered to stimulate the production of PGs through α2-adrenoceptors (Levine et al., 1986). ACh can be released from sensory nerve fibres (Tata et al., 1994), damaged endothelial cells, platelets and parasympathetic postganglionic neurons. ACh can produce burning pain when applied to human skin (Rukwied and Heyer, 1999) by activating sensory C nerve fibres (Steen and Reeh, 1993) through nicotinic receptors (Jinks and Carstens, 1999; Bernardini et al., 2001).
During inflammation, NGF is markedly up-regulated in nerve fibres associated with the inflamed area (Steinhoff et al., 2003) and estrogen also increases endometrial stromal NGF protein expression by 43% (Krizsan-Agbas et al., 2003). NGF can increase the synthesis of SP and CGRP and also directly stimulate degranulation of mast cells (Schaible et al., 2005). NGF can also sensitize (lower the threshold) or excite the terminals of sensory nerve fibres by interacting with cell-surface receptors expressed by these neurons (Julius et al., 2001). Hence, multiple potential molecular mechanisms exist within the cells of endometriotic lesions to cause pain sensation by sensitizing and/or activating sensory nerve fibres.
It will be important to see how future studies reveal variations in these potential mechanisms in endometriosis women who present with pain compared with those who have no complaint of pain, or those who present solely with infertility.
The greatly increased expression of NGF and NGFRp75 by endometriosis lesions may also play a role in inducing ingrowth of nerve fibres into endometriotic tissue in the first place. This expression may play a primary role in setting up the mechanisms for generation of pain from endometriotic lesions.
In summary, human peritoneal endometriotic lesions were innervated by sensory, cholinergic and adrenergic nerve fibres, and sensory and adrenergic nerve fibres were often co-localized. These nerve fibres may interact with each other and with many active molecules released by endometriotic lesions to cause pain and local tenderness. These mechanisms require much more detailed study.
Acknowledgements
This study was supported by research funding from the Department of Obstetrics and Gynaecology, University of Sydney. The authors thank Lawrence Young (Dako, Australia) for technical support.

{kind=link}
{kind=link}