






Abstract
Primary human cytomegalovirus (HCMV) infection during pregnancy is the major cause of congenital viral sequelae. The HCMV-specific T-cell response may have a role in the prevention of virus transmission to the fetus.
HCMV-specific memory T cells were investigated in the second month after primary infection onset in 44 pregnant women (15 transmitting the infection to the fetus) and 8 pregnant women with remote infection. Peripheral blood mononuclear cells were stimulated for 12 days with peptide pools of HCMV proteins IE-1, IE-2, and pp65, and subsequently restimulated for 24 hours with the same peptide pools in a cultured enzyme-linked immunospot (ELISPOT) assay.
In pregnant women with primary infection, the cultured ELISPOT assay detected a higher T-cell response to pp65 than to IE-1 or IE-2, whereas in remote infection pp65-, IE-1–, and IE-2–specific T cells were detected at comparable levels. During primary infection, the cultured ELISPOT response was mainly mediated by CD4+ T cells, and was lower than in remote infection. Strikingly, the cultured ELISPOT response to pp65 (but not to IE-1 or IE-2) was significantly higher in nontransmitting mothers. To detect other factors potentially associated with nontransmission, different serological parameters were analyzed. Only immunoglobulin G avidity index was higher in nontransmitting mothers, who showed also a lower DNAemia level. These 2 parameters remained associated with congenital infection in multivariate analysis.
Determination of HCMV-specific T cells by cultured ELISPOT, in pregnant women with primary HCMV infection, in association with avidity index and DNAemia may help to assess the risk of HCMV fetal transmission.
(See the and .)
Human cytomegalovirus (HCMV) is a major cause of congenital viral infection, occurring in 0.5%–2% of pregnancies [1] and in 20% of cases results in sensory and developmental impairment [2]. HCMV transmission only rarely occurs during nonprimary infection [3], whereas primary infection in pregnancy confers a major risk (about 40%) of congenital infection [4]. Thus, a preexisting immune response appears to provide a strong degree of protection from virus transmission to the fetus. As for the humoral response, in one study, women with primary infections who did not transmit the infection to the fetus exhibited earlier development of neutralizing antibodies against the HCMV pentameric complex gH/gL/pUL128L [5]. Also, the early development of an HCMV-specific CD4+ T-cell lymphoproliferative response [6, 7] has been reported in nontransmitting women. In addition, reduced reexpression of CD45RA, on the surface of HCMV-specific effector memory T cells, correlates with HCMV transmission to the fetus [8, 9]. Studies on transplant recipients have also documented the importance of the HCMV-specific CD4+ T-cell response for the control of viral infection [10–13].
In recent years, the simple tools Quantiferon and enzyme-linked immunospot (ELISPOT) assay were largely used to detect antigen-specific T-cell responses. Using Quantiferon, Eldar-Yedidia and colleagues demonstrated an association between high interferon-gamma (IFN-γ) relative response to HCMV and high risk of transmission to the fetus [14]; in addition, a higher response detected by the ELISPOT assay was shown to be associated with an increased risk of congenital infection [15]. According to the size of the peptides used in both studies, the response was most likely sustained by HCMV-specific CD8+ T cells. Both Quantiferon and standard ELISPOT assays quantify T cells capable of prompt IFN-γ production. These cells are thought to be mainly effector or effector-memory T cells [16]. However, the ELISPOT assay can also be used to quantify antigen-specific T cells with proliferative capacity [17]. In this assay, antigen-specific lymphocytes are expanded for 12 days and then restimulated with the same antigens (cultured ELISPOT). These expandable antigen-specific T cells are thought to be central memory T cells [18]. The aim of this study was to investigate the HCMV-specific memory T-cell response with the cultured ELISPOT assay in pregnant women with primary HCMV infection and evaluate its correlation with HCMV transmission to the fetus, in parallel with the antibody response and viral load.
METHODS
Blood Samples
Forty-four pregnant women experiencing primary HCMV infection during the first or second trimester of gestation were enrolled in the study (Table 1). Heparinized blood samples were collected at a median estimated time of 45 (range, 26–66) days after onset of infection. In addition, 8 pregnant women (median age, 32 [range, 25–36] years) with remote infection (>5 years) were enrolled. This retrospective study was performed according to the guidelines of the Institutional Review Board of Fondazione IRCCS Policlinico San Matteo on the use of biological specimens for scientific purposes in keeping with Italian legislation (art.13 D.Lgs 196/2003).
Characteristics of the 44 Primary Human Cytomegalovirus–Infected Pregnant Women
| Parameters | Nontransmitters (n = 29) | Transmitters (n = 15) | P Value |
|---|---|---|---|
| Age, y, median (range) | 33 (23–40) | 35 (27–41) | NS |
| Gestational week at onset of maternal infection, median (range) | 15 (1–26) | 18 (4–25) | NS |
| Estimated days after onset of infection, median (range) | 47 (26–66) | 43 (29–65) | NS |
| Parameters used for dating infection onset, No. (%) of cases | |||
| Presence of symptoms and/or biochemical signs | 22 (67) | 14 (93) | NS |
| IgG seroconversion | 19 (65) | 10 (66) | NS |
| Low IgG avidity index | 12 (41) | 4 (27) | NS |
| IgM ratio | 7 (24) | 3 (20) | NS |
| Neutralizing antibody seroconversion | 20 (67) | 10 (67) | NS |
| Diagnosis of congenital infection, No. (%) of cases | |||
| At amniocentesis | 13 (45) | 5 (33) | NS |
| At birth | 29 (100) | 14a (93) | NS |
| Infant outcome at birth, No. (%) of cases | |||
| Symptomatic | NA | 1 (7) | NA |
| Asymptomatic | NA | 11 (73) | NA |
| Unknown | NA | 3 (20) | NA |
| Parameters | Nontransmitters (n = 29) | Transmitters (n = 15) | P Value |
|---|---|---|---|
| Age, y, median (range) | 33 (23–40) | 35 (27–41) | NS |
| Gestational week at onset of maternal infection, median (range) | 15 (1–26) | 18 (4–25) | NS |
| Estimated days after onset of infection, median (range) | 47 (26–66) | 43 (29–65) | NS |
| Parameters used for dating infection onset, No. (%) of cases | |||
| Presence of symptoms and/or biochemical signs | 22 (67) | 14 (93) | NS |
| IgG seroconversion | 19 (65) | 10 (66) | NS |
| Low IgG avidity index | 12 (41) | 4 (27) | NS |
| IgM ratio | 7 (24) | 3 (20) | NS |
| Neutralizing antibody seroconversion | 20 (67) | 10 (67) | NS |
| Diagnosis of congenital infection, No. (%) of cases | |||
| At amniocentesis | 13 (45) | 5 (33) | NS |
| At birth | 29 (100) | 14a (93) | NS |
| Infant outcome at birth, No. (%) of cases | |||
| Symptomatic | NA | 1 (7) | NA |
| Asymptomatic | NA | 11 (73) | NA |
| Unknown | NA | 3 (20) | NA |
Abbreviations: IgG, immunoglobulin G; IgM, immunoglobulin M; NA, not applicable; NS, not significant.
aOne termination of pregnancy after amniocentesis.
Characteristics of the 44 Primary Human Cytomegalovirus–Infected Pregnant Women
| Parameters | Nontransmitters (n = 29) | Transmitters (n = 15) | P Value |
|---|---|---|---|
| Age, y, median (range) | 33 (23–40) | 35 (27–41) | NS |
| Gestational week at onset of maternal infection, median (range) | 15 (1–26) | 18 (4–25) | NS |
| Estimated days after onset of infection, median (range) | 47 (26–66) | 43 (29–65) | NS |
| Parameters used for dating infection onset, No. (%) of cases | |||
| Presence of symptoms and/or biochemical signs | 22 (67) | 14 (93) | NS |
| IgG seroconversion | 19 (65) | 10 (66) | NS |
| Low IgG avidity index | 12 (41) | 4 (27) | NS |
| IgM ratio | 7 (24) | 3 (20) | NS |
| Neutralizing antibody seroconversion | 20 (67) | 10 (67) | NS |
| Diagnosis of congenital infection, No. (%) of cases | |||
| At amniocentesis | 13 (45) | 5 (33) | NS |
| At birth | 29 (100) | 14a (93) | NS |
| Infant outcome at birth, No. (%) of cases | |||
| Symptomatic | NA | 1 (7) | NA |
| Asymptomatic | NA | 11 (73) | NA |
| Unknown | NA | 3 (20) | NA |
| Parameters | Nontransmitters (n = 29) | Transmitters (n = 15) | P Value |
|---|---|---|---|
| Age, y, median (range) | 33 (23–40) | 35 (27–41) | NS |
| Gestational week at onset of maternal infection, median (range) | 15 (1–26) | 18 (4–25) | NS |
| Estimated days after onset of infection, median (range) | 47 (26–66) | 43 (29–65) | NS |
| Parameters used for dating infection onset, No. (%) of cases | |||
| Presence of symptoms and/or biochemical signs | 22 (67) | 14 (93) | NS |
| IgG seroconversion | 19 (65) | 10 (66) | NS |
| Low IgG avidity index | 12 (41) | 4 (27) | NS |
| IgM ratio | 7 (24) | 3 (20) | NS |
| Neutralizing antibody seroconversion | 20 (67) | 10 (67) | NS |
| Diagnosis of congenital infection, No. (%) of cases | |||
| At amniocentesis | 13 (45) | 5 (33) | NS |
| At birth | 29 (100) | 14a (93) | NS |
| Infant outcome at birth, No. (%) of cases | |||
| Symptomatic | NA | 1 (7) | NA |
| Asymptomatic | NA | 11 (73) | NA |
| Unknown | NA | 3 (20) | NA |
Abbreviations: IgG, immunoglobulin G; IgM, immunoglobulin M; NA, not applicable; NS, not significant.
aOne termination of pregnancy after amniocentesis.
Diagnosis and Timing of Primary Infection
Diagnosis of primary HCMV infection was based on 1 or more of the following criteria: HCMV-specific immunoglobulin G (IgG) seroconversion, HCMV-specific immunoglobulin M (IgM) antibody detection, low IgG avidity index (AI), and HCMV DNAemia. HCMV-specific IgG and IgM were determined by ETI-CYTOK-G and ETI-CYTOK-M (DiaSorin). IgM results obtained by the commercial assay were confirmed by an in-house–developed capture enzyme-linked immunosorbent assay (ELISA) [19]. IgG AI was determined by an in-house–developed ELISA test using HCMV nuclear antigen [20]. HCMV DNA was determined by real-time polymerase chain reaction [21, 22]. Estimated timing of primary infection was based on the following criteria: decreasing levels of HCMV-specific IgM antibody, increasing levels of AI, increasing levels of neutralizing antibodies detected on human embryonic lung fibroblasts (HELF) and the presence of clinical symptoms along with laboratory findings [23, 24]. Additional serological assays were performed to analyze the possible correlation between the antibody response to HCMV and virus transmission to the fetus. The neutralization assay was performed also in epithelial (ARPE-19) cells [25] and the IgG antibody titers to gH/gL/pUL128L, gH/gL/gO, and gB were determined by ELISA, as reported [5].
Diagnosis of Congenital Human Cytomegalovirus Infection
Congenital HCMV infection was diagnosed before birth by HCMV isolation and/or viral DNA detection in amniotic fluid or fetal blood [26] or within 2 weeks after birth by viral DNA detection in urine.
Isolation of Peripheral Blood Mononuclear Cells
Peripheral blood mononuclear cells (PBMCs) were isolated by standard density gradient centrifugation using Lymphoprep (Axis-Shield). Isolated PBMCs were cryopreserved in RPMI-1640 (Euroclone) supplemented with 10% dimethyl sulfoxide (Sigma-Aldrich) and 5% human albumin (Grifold Biologicals) and preserved in liquid nitrogen.
Cultured Enzyme-Linked Immunospot Assay
PBMCs were thawed, washed, and resuspended in culture medium (RPMI-1640 medium supplemented with 2 mM l-glutamine, 100 U/mL penicillin, 100 µg/mL streptomycin, and 10% heat-inactivated fetal calf serum, Euroclone). Cells were kept overnight at 37°C in a humidified 5% CO2 atmosphere. PBMCs were then transferred to a 48-well flat-bottom plate (5 × 105 cells/mL per well) and stimulated with (i) peptide pools (15-mer overlapping by 11 aa) encompassing the HCMV proteins immediate early (IE) -1, IE-2, and the phosphoprotein (pp) 65 (JPT Peptide Technologies) at a final concentration of 0.25 µg/mL; (ii) culture medium only; (iii) or phytohemagglutinin (PHA; 5 µg/mL, Sigma-Aldrich). PBMCs were then incubated at 37°C in a humidified 5% CO2 atmosphere for 12 days. On days 4 and 8, half of the supernatant from each well was removed and replaced with fresh culture medium supplemented with 20 IU/mL recombinant human interleukin 2 (Peprotech). After 12 days, cells were harvested, washed 3 times with culture medium, and resuspended at a concentration of 4 × 105 cells/mL before their use in the ELISPOT assay. The human IFN-γ ELISPOT kit (Diaclone) and Multiscreen-IP 96-well plates (Merck Millipore) were used as described elsewhere [17]. Cells were added to the plate in triplicate (4 × 104 cells/well) and restimulated for 24 hours with the corresponding peptide pool (0.25 µg/mL) used for the 12-day expansion or staphylococcal enterotoxin B (SEB, 2 µg/mL, Sigma-Aldrich) for cells expanded with PHA. Spots were counted using an automated AID ELISPOT reader system (AutoImmun Diagnostika GmbH). Results are presented as net spots per million PBMCs calculated according to the following formula: (mean number of spots/million PBMCs with antigen) – (mean number of spots/million PBMCs with culture medium).
Cytokine Flow Cytometry
Based on the number of PBMCs available, 9 samples were also analyzed by flow cytometry. After expansion, 0.2–0.4 × 106 cells were transferred to a 96-well round-bottom plate in 40 µL culture medium without fetal calf serum, in the presence of the corresponding peptide pool or SEB or culture medium alone. Following 1 hour incubation at 37°C in a humidified 5% CO2, 140 µL of culture medium supplemented with 10% heat-inactivated fetal calf serum containing brefeldin A (Sigma-Aldrich) at a final concentration of 10 µg/mL was added. After overnight incubation, cells were washed in phosphate-buffered saline (PBS) ethylenediaminetetraacetic acid 2mM and incubated with the Live/Dead Fixable Far Red dead cell stain kit (Thermo Fisher Scientific) for 30 minutes at 4°C. Cells were then washed with PBS, fixed, and permeabilized with BD Cytofix/Cytoperm (BD Biosciences) according to the manufacturer’s instructions for intracellular staining with the following monoclonal antibodies: CD3 PC5, CD4 ECD, CD8 PC7, IFN-γ FITC (all from Beckman Coulter). Finally, cells were resuspended in 1% paraformaldehyde and analyzed with a Navios flow cytometer (Beckman Coulter).
Statistical Analysis
Categorical variables are described as count and percentage. Fisher exact test was used to compare the difference in frequencies. Quantitative variables are described as median and range; comparisons between primary and remote infections or between HCMV transmitting and nontransmitting women were made with Mann-Whitney U test. Receiver operating characteristic (ROC) analysis was performed to establish the best cutoff to identify transmitting and nontransmitting women. (Prism 6 software, GraphPad Software). Furthermore, in addition to cultured ELISPOT responses, different parameters were considered in relation to the transmission of HCMV to the fetus (DNAemia, AI, IgG directed to gB, gH/gL/gO, and the pentameric complex gH/gL/pUL128L and antibodies neutralizing the infection of epithelial cells ARPE-19 and HELF). Parameters showing different levels in transmitting and nontransmitting women (with a P value <.10) were subsequently analyzed in a multivariate logistic regression model; P < .05 was considered statistically significant. All tests were 2-sided. The data analysis was performed with the Stata statistical package (release 14.0, 2015, StataCorp LLC) and NCSS (release 2006, NCSS LLC).
RESULTS
Human Cytomegalovirus-Specific T-Cell Response in Immunocompetent Pregnant Women Measured by Cultured Enzyme-Linked Immunospot
Memory T cells with proliferative capacity directed against pp65, IE-1, and IE-2 were measured in 44 pregnant women in the second month after onset of primary HCMV infection and in 8 pregnant women with remote infection (Figure 1). The median number of net spots/million PBMCs in response to pp65, IE-1, and IE-2 was significantly lower in primary than remote infection. In addition, a significantly higher response to pp65 than to IE-1 (P < .05) and IE-2 (P = .01) was observed in primary infections. No significant difference was observed between primary and remote infections in response to SEB. Flow cytometry analysis showed that, among 9 women with primary infection examined, 6 (67%) had a detectable response against at least 1 of the 3 proteins. Most IFN-γ producing antigen-specific T cells were CD4+ (Figure 2).
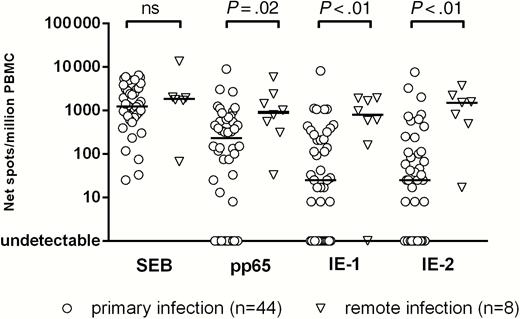
T-cell response to staphylococcal enterotoxin B (SEB) and human cytomegalovirus proteins pp65, IE-1, and IE-2, detected by cultured enzyme-linked immunospot assay, in pregnant women with primary or remote infection. Results are expressed as net spots/million peripheral blood mononuclear cells (PBMCs); P values, determined by Mann-Whitney U test, are reported; Abbreviation: ns, not significant (P > .05).
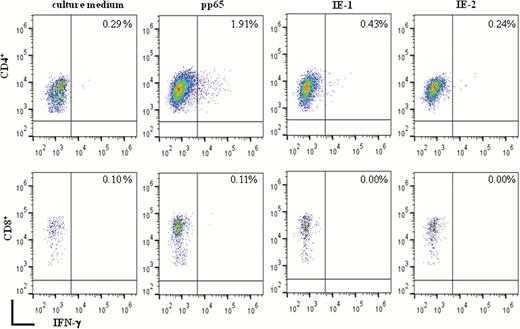
Flow cytometry detection of interferon gamma (IFN-γ) produced by memory T cells with proliferative capacity in the presence of culture medium alone or after stimulation with pp65, IE-1, and IE-2. Percentages of IFN-γ–producing CD4+ and CD8+ T cells examined in a representative pregnant woman with primary human cytomegalovirus infection are reported.
Human Cytomegalovirus-Specific Memory T-Cell Response and Virus Transmission to the Fetus
Memory T cells with proliferative capacity specific for pp65, IE-1, and IE-2 were compared between 15 women who transmitted and 29 women who did not transmit the infection to the fetus (Figure 3). The memory T-cell response specific for pp65 was significantly lower (P < .01) in transmitting (median, 13 [range, 0–1150] spots/million PBMCs) than in nontransmitting mothers (median, 388 [range, 0–8867] spots/million PBMCs). No differences were observed in the response to IE-1 or IE-2, between transmitting and nontransmitting women (both groups showed low number of net spots/million), as well as for the response to SEB.
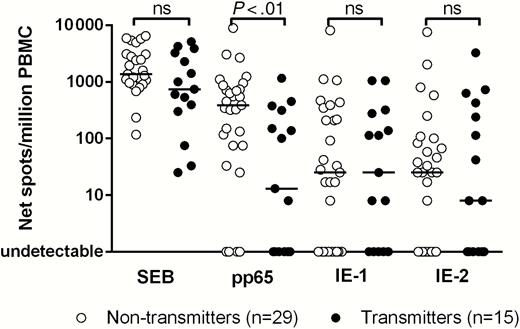
T-cell response to staphylococcal enterotoxin B (SEB) and human cytomegalovirus proteins pp65, IE-1, and IE-2, detected by cultured enzyme-linked immunospot assay in 29 nontransmitting and 15 transmitting women; P values, determined by Mann-Whitney U test, are reported; Abbreviations: ns, not significant (P > .05); PBMC, peripheral blood mononuclear cell.
Receiver Operating Characteristic Analysis and Risk of Human Cytomegalovirus Transmission to the Fetus
ROC analysis showed that a pp65 specific T-cell response of 20 net spots/million PBMCs (Figure 4) was the best cutoff to discriminate between transmitting and nontransmitting women (area under the curve = 0.74, sensitivity 53%, specificity 86%, positive predictive value = 67%, negative predictive value = 78%). Among women with a response ≤20 spots/million, 4 of 12 (33.3%) did not transmit the infection to the fetus, whereas 25 of 32 (78.1%) women with a response >20 spots/million PBMCs did not transmit the infection (P = .01; Figure 5). On the basis of this cutoff, an odds ratio of 7.14 (95% confidence interval, 1.65–30.89) for nontransmission of the virus to the fetus was associated with a pp65-specific T-cell response >20 spots/million PBMCs.
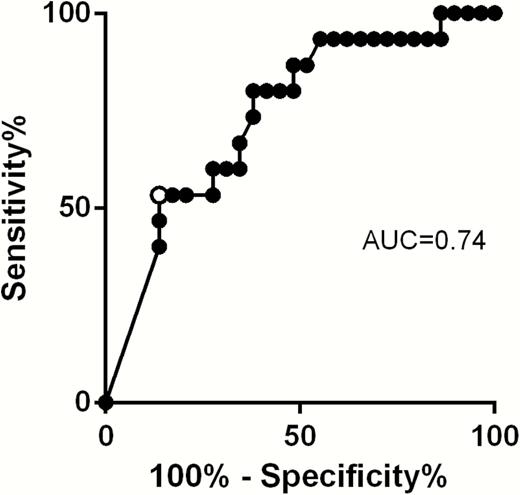
Receiver operating characteristic curve for the establishment of the best cutoff to discriminate transmitting and nontransmitting women. Black dots represent values of cultured enzyme-linked immunospot assay corresponding to different sensitivity and specificity. White dot represents the cutoff of 20 spots/million peripheral blood mononuclear cells. Abbreviation: AUC, area under the curve.
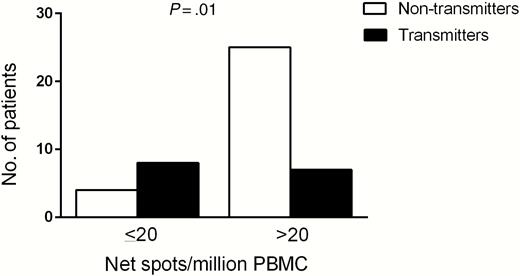
Cultured enzyme-linked immunospot assay response and number of nontransmitting (white histogram) or transmitting (black histogram) women divided in 2 groups: >20 or ≤20 spots/million peripheral blood mononuclear cells (PBMCs); P value, determined by Fisher exact test, is reported.
Immunological and Virological Parameters Associated With Human Cytomegalovirus Transmission to the Fetus
In addition to the T-cell response, we analyzed the possible association of antibody response with virus transmission to the fetus. The kinetics of IgG, IgM, and IgG AI to total HCMV antigen and IgG directed to envelope glycoprotein complexes (gB, gH/gL/gO, gH/gL/pUL128L) and the neutralizing antibody response vs HCMV infection of ARPE-19 and HELF during primary infection was investigated elsewhere [24]. In the 44 pregnant women analyzed in the present study, none of these parameters was found to be significantly different between transmitting and nontransmitting women (data not shown), with the exception of AI—that is, nontransmitting women showed higher levels of the IgG AI to whole virus antigen (P = .06; Figure 6A). Then, we examined DNAemia levels in transmitting and nontransmitting women, and we found that DNAemia levels were significantly lower in nontransmitting women (P = .04; Figure 6B).
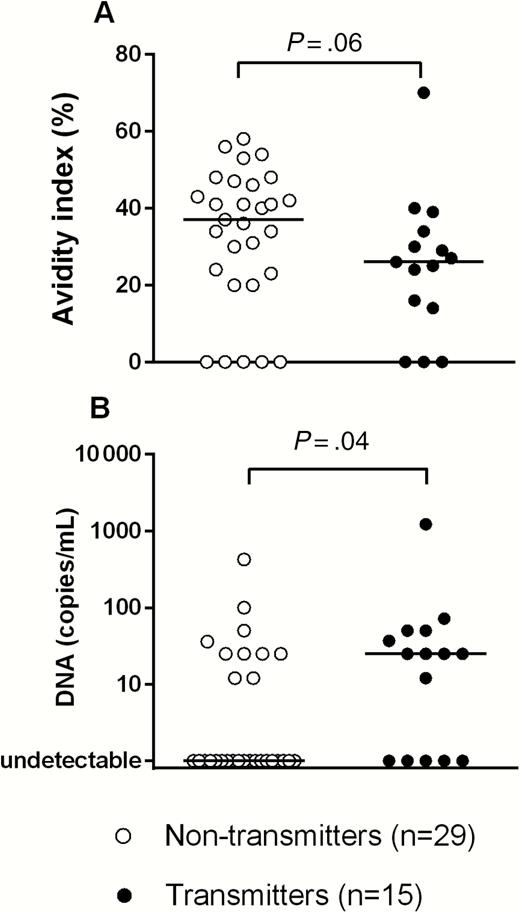
Immunoglobulin G avidity index (A) and DNAemia (B) in 29 nontransmitting and 15 transmitting women. P values, determined by Mann-Whitney U test, are reported.
The 3 parameters detected as significantly different between transmitting and nontransmitting mothers were analyzed in a multivariate logistic regression model, and high cultured ELISPOT response to pp65, high AI, and low DNAemia were independently associated with lack of HCMV transmission (Table 2). However, cultured ELISPOT to pp65 showed the highest association.
Multivariate Logistic Regression Analysis
| Parameters | P Value | Odds Ratio (95% CI) |
|---|---|---|
| pp65 cultured ELISPOT (>20 spots/million PBMCs) | <.01 | 33.15 (2.96–371.96) |
| Avidity index (for each 10% increase) | .03 | 1.93 (1.07–3.47) |
| DNAemia (log10 copies/mL) | .01 | 0.19 (.05–.70) |
| Parameters | P Value | Odds Ratio (95% CI) |
|---|---|---|
| pp65 cultured ELISPOT (>20 spots/million PBMCs) | <.01 | 33.15 (2.96–371.96) |
| Avidity index (for each 10% increase) | .03 | 1.93 (1.07–3.47) |
| DNAemia (log10 copies/mL) | .01 | 0.19 (.05–.70) |
Abbreviations: CI, confidence interval; ELISPOT, enzyme-linked immunospot assay; PBMC, peripheral blood mononuclear cell.
Multivariate Logistic Regression Analysis
| Parameters | P Value | Odds Ratio (95% CI) |
|---|---|---|
| pp65 cultured ELISPOT (>20 spots/million PBMCs) | <.01 | 33.15 (2.96–371.96) |
| Avidity index (for each 10% increase) | .03 | 1.93 (1.07–3.47) |
| DNAemia (log10 copies/mL) | .01 | 0.19 (.05–.70) |
| Parameters | P Value | Odds Ratio (95% CI) |
|---|---|---|
| pp65 cultured ELISPOT (>20 spots/million PBMCs) | <.01 | 33.15 (2.96–371.96) |
| Avidity index (for each 10% increase) | .03 | 1.93 (1.07–3.47) |
| DNAemia (log10 copies/mL) | .01 | 0.19 (.05–.70) |
Abbreviations: CI, confidence interval; ELISPOT, enzyme-linked immunospot assay; PBMC, peripheral blood mononuclear cell.
DISCUSSION
In this study, memory HCMV-specific T cells with proliferative capacity were investigated in pregnant women experiencing primary HCMV infection. We used a cultured ELISPOT and 15-mer peptide pools that allow the detection of CD4+ and CD8+ T cells, regardless of human leukocyte antigen [27, 28]. At 1–2 months after infection onset, specific T cells detected by the cultured ELISPOT (mostly CD4+ T cells) were mainly directed to the HCMV pp65. A higher cultured ELISPOT response, associated with a higher AI and low DNAemia, was associated with a lower risk of transmission to the fetus.
While the standard ELISPOT allows detection of effector or effector-memory T cells, the cultured ELISPOT (through antigen-specific expansion of T cells for 12 days) detects memory T cells with proliferative capacity [17, 29]; these cells display characteristics of central memory T cells [18]. Pregnant women with remote infection exhibited memory T cells with proliferative capacity specific for pp65, IE-1, and IE-2. Pregnant women examined in the second month after onset of primary HCMV infection had a significantly lower response to the 3 proteins examined with respect to remote infections, and the pp65-specific T-cell response was significantly greater with respect to IE-1 and IE-2. This may be due to the fact that the IE-specific T cells are mainly CD8+, and this subset is poorly expandable in vitro during primary infection [7]. On the contrary, CD4+ T cells are the major responders in the cultured ELISPOT during primary infection and recognize pp65 better than IE antigens. This response is different from that observed after T-cell stimulation for a few hours. With the latter assay, effector-memory CD4+ and CD8+ T cells specific for pp65 and IE-1 are already detectable within a few weeks after infection onset [30]. While effector T cells, which promptly limit the infection, have a lower proliferative potential in vitro (thus, are not detected by the cultured ELISPOT), memory T cells with proliferative capacity appear much later.
By taking advantage of the cultured ELISPOT assay, we observed a significantly higher proliferative T-cell response (to pp65 but not to IE-1 or IE-2) in nontransmitting women. Conversely, a recent study adopting the standard ELISPOT reported apparently discordant results, as higher T-cell response, most likely CD8+, was associated with HCMV transmission to the fetus [15]. However, as mentioned above, the standard ELISPOT detects effector T cells (regardless of their proliferative capacity). In addition, in the aforementioned study, primary and remote infections were grouped together. When considering only primary infections, we found no significant difference for CD4+ and CD8+ IE-1– or pp65-specific effector T-cell responses (detected by flow cytometry) between transmitting and nontransmitting women [31]. Instead, the more rapid development of a particular effector memory T-cell subset, reexpressing CD45RA (TEMRA), was associated with the control of infection [8, 9, 32]. HCMV-specific T-cell responses in women transmitting the infection was also examined by Quantiferon [14]. In this case, the amount of IFN-γ released by HCMV-specific CD8+ T cells (normalized on the IFN-γ response to PHA) was lower in nontransmitting women. As it was observed that transmitting women had a delayed clearance of HCMV DNA in blood [33], it is possible that a strong, specific T-cell response is induced by higher amounts of antigen. In these previous studies, only the “effector-like” response (IFN-γ production) of HCMV-specific T cells was determined, regardless of their “memory-like” response (ie, proliferation). In the present study, we investigated a particular subset of T cells that are important for the establishment of long-term memory. Thus, it seems that higher levels of the effector T-cell response and lower levels of T cells with proliferative capacity are associated with transmission to the fetus.
During primary infection, HCMV-specific T cells have a decreased ability to proliferate and secrete interleukin 2 [6, 7, 34]. Most important, and in agreement with the present results, a delayed HCMV-specific CD4+ lymphoproliferative response was observed in women transmitting the infection to the fetus [6, 7]. In the present study, we observed that memory T cells with proliferative capacity detected by cultured ELISPOT within 2 months after onset of infection were mainly CD4+. The crucial role of maternal CD4+ T cells in preventing CMV transmission to the fetus is supported by a study of Bialas and colleagues [35], in which the depletion of CD4+ T cells in the rhesus macaque model was associated with a reduced virus-specific CD8+ T-cell response, delayed production of neutralizing antibodies, and higher plasma and amniotic fluid viral load. Additionally, in transplant recipients who do not spontaneously control HCMV infection, significantly lower levels of CD4+ T cells were observed [12].
In addition to the high proliferative T-cell response, we observed that high IgG AI was associated with a lower risk of transmission to the fetus. The appearance of early antibody with high avidity was previously correlated with a lower risk of HCMV transmission to the fetus [36]. On the contrary, neither neutralization titer nor IgG to glycoprotein complexes was found to be associated with transmission in the second month after infection, although in a previous study we found higher titers of IgG to gH/gL/gO and gH/gL/pUL128L in nontransmitting mothers at an earlier time-point [5]. Finally, as already reported, a lower DNAemia was detected in nontransmitting mothers [5, 33]. Multivariate logistic regression analysis showed that the 3 parameters (high proliferative T-cell response, high AI, and low DNAemia) were all independently associated with a lower risk of congenital infection, although the T-cell response showed the highest association.
Our observation that both T-cell and antibody response correlate with the outcome of maternal primary infection may suggest that an ideal candidate vaccine should be able to elicit both humoral and T-cell response against HCMV. Accordingly, in a recent study in the guinea pig model, the combined vaccination with gB and pp65 was able to induce both humoral and cellular response and conferred better protection against maternal viremia than monovalent vaccines [37].
Limitations of our study are the small number of patients examined and the adoption of nonstandardized techniques. In particular, it should be verified also whether the cultured ELISPOT assay could be exported outside the research setting. However, results of the logistic regression analysis indicate that the combination of different parameters (T-cell response, serology, and DNAemia) could provide useful information for the assessment of risk of HCMV congenital infection. In conclusion, confirmation of results reported in a larger population should be achievable in the next future.
Notes
Acknowledgments. We thank Daniela Sartori for editing the manuscript; Laurene Kelly for revision of the English; and Daniele Lilleri for helpful discussion and revision of the manuscript.
Financial support. This work was supported by the Ministero della Salute, Ricerca Corrente (grant number 80207) and Fondazione Cariplo (grant number 2012-0626).
Potential conflicts of interest. All authors: No reported conflicts of interest. All authors have submitted the ICMJE Form for Disclosure of Potential Conflicts of Interest. Conflicts that the editors consider relevant to the content of the manuscript have been disclosed.
References


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Comments