






Abstract
This review examines the association of a subset of endothelial nitric oxide synthase gene (NOS3) polymorphisms (Glu298Asp, intron 4, and -786T>C) with cardiovascular disease. The Glu298Asp polymorphism within exon 7 is the only common nonsynonymous variant. The variants have been associated with low plasma nitric oxide concentrations and reduced vascular reactivity; difficulties in measuring those phenotypes means that their functional role remains unclear. A large meta-analysis of NOS3 polymorphisms in coronary heart disease revealed per-allele odds ratios of 1.17 (95% confidence interval: 1.07, 1.28) for Glu298Asp, 1.17 (95% confidence interval: 1.07, 1.28) for -786T>C, and 1.12 (95% confidence interval: 1.01, 1.24) for intron 4. However, there was evidence that small studies with more striking results could affect the associations of the Glu298Asp and -786T>C polymorphisms with coronary heart disease. Associations of NOS3 polymorphisms with hypertension, preeclampsia, stroke, and diabetes remain uncertain. To date, no reliable gene-gene or gene-environmental interactions have been described. Use of these variants in predictive testing is unlikely to be useful, although the population attributable fraction could be substantial if the modest associations are causal. The need for large-scale genetic association studies using tagging polymorphisms is warranted to confirm or refute a role of the NOS3 gene in coronary heart disease.
GENE
Endothelial nitric oxide synthase (eNOS) is one of three isoforms of nitric oxide synthase that exhibits homology of sequence and function (1). The NOS3 gene was cloned in 1993 and was localized to chromosome 7q35-36 (2). Spanning 4.4 kb of genomic DNA, the gene comprises 26 exons that encode a 135-kD protein containing 1,203 amino acids. Approximately 1,500 base pairs of upstream promoter sequence have also been characterized and contain transcription factor-binding sites that mediate regulation by shear stress and estrogens, among others (3). The eNOS protein synthesizes nitric oxide constitutively via a reaction including the conversion of L-arginine to L-citrulline, which involves the transfer of five electrons provided by nicotinamide adenine dinucleotide phosphate (4). The enzyme acts as a homodimer that can be divided functionally into two major domains: a C-terminal reductase domain and an N-terminal oxygenase domain (5). Catalytic activity requires the presence of heme and the cofactors tetrahydrobiopterin, flavin adenine dinucleotide, flavin mononucleotide, and calmodulin (5). Nitric oxide is not stored but rather released upon its synthesis. Thus, nitric oxide generation is regulated through alterations in the expression or activity of the eNOS enzyme itself or through changes in the availability of activating cofactors or endogenous inhibitor molecules (6, 7).
Nitric oxide from the endothelium is considered an important atheroprotective mediator, and acquired defects in generation of nitric oxide are associated with increases in cardiovascular risk factors (8). Endothelium-dependent, flow-mediated dilatation of the brachial artery (a largely nitric oxide–dependent response) is impaired in young, healthy individuals with a first-degree relative who died from coronary heart disease (CHD) before age 55 years when compared with age-matched individuals with no family history of CHD (9, 10). In addition, mice in which the NOS3 gene has been deleted are hypertensive, and those with deletions in both the apolipoprotein E and NOS3 genes have increased susceptibility to atherosclerosis (11). Because endothelial nitric oxide availability is regulated at the level of synthesis, the gene that encodes eNOS is a candidate for cardiovascular disease (3).
GENE VARIANTS
The NOS3 gene has been extensively screened for variation. Variants detected include numerous single nucleotide polymorphisms (SNPs) (12–14), a variable-number tandem repeat in the intron 4 (15), and a CA repeat microsatellite marker in the intron 13 (12). The only common variation identified that leads to an amino acid substitution in the mature protein is the G894T or Glu298Asp (rs1799983) variant, in which a guanine→thymine substitution at exon 7 leads to a glutamate→aspartate substitution at position 298 (12). Several promoter SNPs have been identified, but there is no clear evidence that any of them lies directly within the consensus sequence for a known transcription factor of NOS3. Similarly, no variations in the 3′ untranslated region have been reported (14). Variation in this region might influence RNA stability (3).
Web appendix tables 1, 2, and 3 describe genotype frequencies in apparently healthy subjects from 64 sample populations, divided according to ethnic background. (This information is presented in the first three of six supplementary tables; each is referred to as “Web appendix table” in the text and is posted on the website of the Human Genome Epidemiology Network (http://www.cdc.gov/genomics/hugenet/reviews.htm) as well as on the Journal's website (http://aje.oupjournals.org/). This review also includes Web Appendix text and 10 supplementary Web figures; each of these figures is referred to as “Web appendix figure” and is also posted online.) A significant difference in the frequency of Asp298 and -786C alleles by ethnic group has been reported previously (16) and was confirmed in a previous meta-analysis of NOS3 genotype and CHD, in which a lower frequency of homozygosity for the Asp298 and -786C alleles was observed among Asians (Asp/Asp—Asians: 0.48 percent vs. non-Asians: 10.73 percent; C/C—Asians: 7.6 percent vs. non-Asians: 32.3 percent) (17). The proportion of subjects homozygous for the intron 4 a allele was similar among Asians and non-Asians (1.6 percent and. 2.0 percent, respectively) (17). A low frequency of subjects homozygous for the Asp298 allele has been reported among Amerindians and mixed Hispanic populations (18–20), which means that very large sample sizes would be needed to obtain reliable estimates of the effect of these polymorphisms in these populations.
Functional variation in the NOS3 gene has yet to be completely characterized. Much attention has focused on three putatively functional variants (-786T>C (rs2070744), intron 4 27-base-pair repeat, and Glu298Asp (rs1799983)), but little information has been available as to how these variants associate with one another. Focusing research efforts on the three variants examined to date limits the study of NOS3 to an isolated “candidate polymorphism” rather than a “candidate gene” approach (21). Knowledge of haplotypes and linkage disequilibrium patterning through the NOS3 gene would enable a more thorough investigation of the role of NOS3 in the development of cardiovascular disease.
We examined the association between the three commonly studied variations in a sample of 2,266 males of British descent from the Northwick Park Heart Study-II. The characteristics of this population-based prospective cohort study have been described elsewhere (22). Haplotypes generated from these three variants and corresponding linkage disequilibrium values are shown in Web appendix tables 4 and 5. The loose association between these three variants, as shown by pairwise (r2) values, justifies direct genotyping of each variant.
Investigation of NOS3 variation was then expanded by studying the International HapMap Project and the University of Washington Variation Discovery Resource (Seattle SNPs) (14, 23). The goal of the HapMap project is to provide SNP data across the entire genome with an average density of one SNP every 1 kb, a resource invaluable for haplotype-based association studies (23). The Seattle SNPs project focuses on characterizing variation across the entire length of specific genes associated with inflammation and cardiovascular disease. We used these data to characterize the pattern of linkage disequilibrium in and around the NOS3 locus in northern European populations. First, we examined linkage disequilibrium across a 110-kb region containing the NOS3 gene by using 21 SNPs from the HapMap database (23). We then focused specifically on the 25 kb of the NOS3 gene itself by using the University of Washington Variation Discovery Resource data (14).
The HapMap data showed the NOS3 gene to be located at the edge of a region of elevated linkage disequilibrium that extends at least 45 kb upstream of the gene, while linkage disequilibrium downstream of the NOS3 gene breaks down abruptly (Web appendix figure 1). An examination of fine-scale linkage disequilibrium across the gene itself, using genotype data from the complete resequencing of the gene by the Seattle SNPs project, confirmed the pattern of elevated linkage disequilibrium across the gene depicted in the gross-scale analysis.
We selected tagging SNPs (tSNPs) for NOS3 based on haplotypes inferred from the Seattle SNPs data by using only those variants with a minor allele frequency of greater than 5 percent. We used the haplotype r2 method (24–26) and applied a minimum coefficient of determination of 0.80 in predicting the state of each tagged SNP. To combine a tagging and functional approach, we included the putatively functional variants -786T>C (rs2070744) and Glu298Asp (rs1799983) as tSNPs regardless of their coefficients. Unfortunately, the intron 4 27-base-pair repeat was not genotyped. The following set of six tSNPs satisfied these conditions: 1) tSNP1: rs2070744 (-786T>C); 2) tSNP2: rs3918167; 3) tSNP3: rs1799983 (Glu298Asp); 4) tSNP4: any one of rs3918188, rs3918181, rs3918182, or rs3918184; 5) tSNP5: any one of rs743506, rs743507, or rs2256314; and 6) tSNP6: rs11539284.
Genotyping of these six variants in a population of northwest European descent will not only directly examine the -786T>C and Glu298Asp variants but also allow assessment of all common variation across the NOS3 gene, with minimal loss of power compared with genotyping all variants directly. Haplotypes generated by these tSNPs represent the common haplotypes in populations of northwest European origin (table 1).
rs2070744 | rs3918167 | rs1799983 | rs3918184 | rs743506 | rs11539284 | Frequency (%) |
|---|---|---|---|---|---|---|
| C | A | T | C | G | G | 0.26 |
| C | G | G | C | A | T | 0.17 |
| T | A | T | C | A | G | 0.12 |
| T | A | G | T | A | T | 0.11 |
| C | A | G | C | A | G | 0.06 |
| T | A | G | T | A | G | 0.05 |
| T | A | T | C | A | T | 0.05 |
| T | A | G | C | G | G | 0.05 |
| T | A | G | C | A | G | 0.04 |
| C | A | T | C | A | T | 0.04 |
| C | A | T | C | A | G | 0.03 |
| C | A | G | T | A | G | 0.02 |
rs2070744 | rs3918167 | rs1799983 | rs3918184 | rs743506 | rs11539284 | Frequency (%) |
|---|---|---|---|---|---|---|
| C | A | T | C | G | G | 0.26 |
| C | G | G | C | A | T | 0.17 |
| T | A | T | C | A | G | 0.12 |
| T | A | G | T | A | T | 0.11 |
| C | A | G | C | A | G | 0.06 |
| T | A | G | T | A | G | 0.05 |
| T | A | T | C | A | T | 0.05 |
| T | A | G | C | G | G | 0.05 |
| T | A | G | C | A | G | 0.04 |
| C | A | T | C | A | T | 0.04 |
| C | A | T | C | A | G | 0.03 |
| C | A | G | T | A | G | 0.02 |
Polymorphism rs2070744 corresponds to the -786T>C variant, and rs179983 corresponds to the Glu298Asp variant.
SeattleSNPs. Program for Genomic Applications. Supported by the National Heart, Lung, and Blood Institute, Seattle, WA. (URL: http://pga.gs.washington.edu). (Accessed January 1, 2005).
rs2070744 | rs3918167 | rs1799983 | rs3918184 | rs743506 | rs11539284 | Frequency (%) |
|---|---|---|---|---|---|---|
| C | A | T | C | G | G | 0.26 |
| C | G | G | C | A | T | 0.17 |
| T | A | T | C | A | G | 0.12 |
| T | A | G | T | A | T | 0.11 |
| C | A | G | C | A | G | 0.06 |
| T | A | G | T | A | G | 0.05 |
| T | A | T | C | A | T | 0.05 |
| T | A | G | C | G | G | 0.05 |
| T | A | G | C | A | G | 0.04 |
| C | A | T | C | A | T | 0.04 |
| C | A | T | C | A | G | 0.03 |
| C | A | G | T | A | G | 0.02 |
rs2070744 | rs3918167 | rs1799983 | rs3918184 | rs743506 | rs11539284 | Frequency (%) |
|---|---|---|---|---|---|---|
| C | A | T | C | G | G | 0.26 |
| C | G | G | C | A | T | 0.17 |
| T | A | T | C | A | G | 0.12 |
| T | A | G | T | A | T | 0.11 |
| C | A | G | C | A | G | 0.06 |
| T | A | G | T | A | G | 0.05 |
| T | A | T | C | A | T | 0.05 |
| T | A | G | C | G | G | 0.05 |
| T | A | G | C | A | G | 0.04 |
| C | A | T | C | A | T | 0.04 |
| C | A | T | C | A | G | 0.03 |
| C | A | G | T | A | G | 0.02 |
Polymorphism rs2070744 corresponds to the -786T>C variant, and rs179983 corresponds to the Glu298Asp variant.
SeattleSNPs. Program for Genomic Applications. Supported by the National Heart, Lung, and Blood Institute, Seattle, WA. (URL: http://pga.gs.washington.edu). (Accessed January 1, 2005).
FUNCTION
Glu298Asp
Some mechanistic studies have been published suggesting a functional effect of the Glu298Asp polymorphism. Associations have been described between the Glu298Asp polymorphism and nitric oxide synthesis (27, 28) and endothelial function (29, 30). A mechanism by which eNOS Asp298 might reduce nitric oxide bioavailability has also been reported (27, 28). In terms of enzymatic activity, studies of recombinant eNOS Asp298 and Glu298 showed no discernible difference in the Michaelis constant Km, nor the Vmax, of the two forms of the enzyme (3, 31, 32). Moreover, there was no difference in Ki for the endogenous methylarginine inhibitors of eNOS, namely, asymmetric dimethylarginine and NG monomethyl-L-arginine (3). The Glu298Asp polymorphism lies within a loop on the external surface of the structure and does not make contact with either the active site of the enzyme or the dimerization interface (3), suggesting that, if functional, the polymorphism would have to exert its effect by a mechanism independent of nitric oxide synthase catalysis. Two studies have recently shown the eNOS protein containing Asp at position 298 to be subject to selective proteolytic cleavage in endothelial cells and vascular tissues (27, 28). If this observation is correct, the cleaved fragments would be expected to lack nitric oxide synthase activity (29, 30). However, two other reports suggest that this observation might be an artifact (32, 33), so further in vitro work is needed to resolve this issue.
Human studies suggest that individuals homozygous for Asp298 experience a reduced blood pressure fall following exercise training (34) and lower basal blood flow and reduced vasodilation to adenosine in their coronary arteries (35). In addition, in some but not all studies (29, 37, 38), subjects homozygous for the Asp298 allele have an enhanced systemic pressor response to phenylephrine (36) and a reduced flow mediated dilatation of the brachial artery. These observations require confirmation in larger studies.
-786T>C
Given the location of -786T>C in the promoter region of NOS3, studies examining the functionality of this polymorphism have focused on eNOS expression levels. Lower eNOS mRNA and serum nitrite/nitrate levels have been found in individuals with the -786C variant (39). Reporter gene assays support such a role (40). Recently, a nuclear protein that exhibits differential binding to the promoter containing the -786T and -786C alleles has also been described (41). Human studies suggest that subjects homozygous for the -786C allele have a decreased maximal forearm blood flow response to acetylcholine, a pharmacologic tool to evaluate nitric oxide production in vivo (42). However, these associations have yet to be reproduced in other functional and population-based studies (22, 43).
Intron 4
Given the intronic location of the intron 4 repeat unit, it is perhaps less likely to be functional. Conflicting associations between the intron 4 variant and nitric oxide pathway activity have been described. Some reports indicate that carriers of this variant have lower nitric oxide plasma levels and decreased protein expression (44, 45), but this finding is not supported by all studies (22, 46). It is possible that the variant is in linkage disequilibrium with other functional variants in regulatory regions of the NOS3 gene.
DISEASES (OUTCOMES)
Information about the epidemiology of CHD, stroke, hypertension, and preeclampsia (47–61) can be found in the online Appendix.
GENE-DISEASE ASSOCIATIONS
CHD
For genetic association studies evaluating the role of the Glu298Asp, -786T>C, or intron 4 polymorphisms of the NOS3 gene in CHD, we conducted an updated meta-analysis of studies in all languages published until February 2006. The search, selection criteria, data abstraction, and statistical methods are described in the online Appendix Methods section (62, 63). Briefly, our principal hypothesis was that an additive (per-allele) model for NOS3 Asp298, -786T>C, or intron 4 a variants would be associated with an increased risk of CHD. Secondary analyses involved recessive, dominant models and pairwise comparisons of the genotype groups generated. For all models used, the minor allele was considered the risk allele.
Of the 71 studies (69 articles) identified (12, 15, 22, 64–128), 64 (62 articles) were included in this updated meta-analysis (12, 15, 22, 64–121). Information on study design, genotype frequencies, patient characteristics, and outcomes description of studies included in the meta-analyses is outlined in Web appendix tables 1, 2, and 3. Four studies were excluded because duplication or partial overlapping was considered likely after contacting the study author (122–125), and three were omitted because relevant data were not reported and could not be obtained from study authors (126–128).
Glu298Asp polymorphism.
The meta-analysis of the Glu298Asp polymorphism included 42 studies (40 articles) comprising 13,876 CHD cases and 13,042 controls (12, 22, 64–74, 86–89, 91–94, 96–99, 102, 103, 106–108, 111–116, 118–121). The odds ratio under an additive model for CHD was 1.17 (95 percent confidence interval (CI): 1.07, 1.28; p = 0.001; Web appendix figure 2). However, there was evidence of substantial between-study heterogeneity (I2 = 67.9 percent,
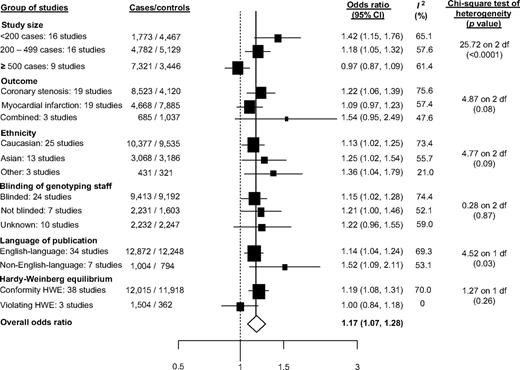
Studies of the Glu298Asp polymorphism and risk of coronary heart disease grouped by study characteristics. The sizes of the boxes relate to the inverse of the variance and thus to study size. CI, confidence interval; HWE, Hardy-Weinberg equilibrium.
Comparison | Glu298Asp | Intron 4 | -786T>C | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Odds ratio | 95% CI§ | Odds ratio | 95% CI | Odds ratio | 95% CI | ||||
| Additive model | |||||||||
| Random | 1.17 | 1.07, 1.28 | 1.12 | 1.01, 1.24 | 1.17 | 1.07, 1.28 | |||
| I2 (p for heterogeneity) | 67.9% (<0.0001) | 55.4% (<0.0001) | 62.7% (<0.0001) | ||||||
| Homozygous for rare allele vs. homozygous for common allele | |||||||||
| Random | 1.36 | 1.12, 1.66 | 1.24 | 0.97, 1.60 | 1.35 | 1.11, 1.64 | |||
| I2 (p for heterogeneity) | 60.1% (<0.0001) | 21.5% (0.148) | 52.6% (0.003) | ||||||
| Heterozygous vs. homozygous for common allele | |||||||||
| Random | 1.07 | 0.98, 1.17 | 1.10 | 0.98, 1.24 | 1.13 | 1.00, 1.26 | |||
| I2 (p for heterogeneity) | 46.4% (0.001) | 53.3% (<0.0001) | 54.2% (0.002) | ||||||
| Recessive model | |||||||||
| Random | 1.34 | 1.11, 1.62 | 1.20 | 0.95, 1.51 | 1.24 | 1.07, 1.47 | |||
| I2 (p for heterogeneity) | 58% (<0.0001) | 14.8% (0.238) | 40.2% (0.03) | ||||||
| Dominant model | |||||||||
| Random | 1.15 | 1.04, 1.27 | 1.12 | 1.00, 1.26 | 1.17 | 1.05, 1.32 | |||
| I2 (p for heterogeneity) | 59.5% (<0.0001) | 55.8% (<0.0001) | 60.2% (<0.0001) | ||||||
Comparison | Glu298Asp | Intron 4 | -786T>C | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Odds ratio | 95% CI§ | Odds ratio | 95% CI | Odds ratio | 95% CI | ||||
| Additive model | |||||||||
| Random | 1.17 | 1.07, 1.28 | 1.12 | 1.01, 1.24 | 1.17 | 1.07, 1.28 | |||
| I2 (p for heterogeneity) | 67.9% (<0.0001) | 55.4% (<0.0001) | 62.7% (<0.0001) | ||||||
| Homozygous for rare allele vs. homozygous for common allele | |||||||||
| Random | 1.36 | 1.12, 1.66 | 1.24 | 0.97, 1.60 | 1.35 | 1.11, 1.64 | |||
| I2 (p for heterogeneity) | 60.1% (<0.0001) | 21.5% (0.148) | 52.6% (0.003) | ||||||
| Heterozygous vs. homozygous for common allele | |||||||||
| Random | 1.07 | 0.98, 1.17 | 1.10 | 0.98, 1.24 | 1.13 | 1.00, 1.26 | |||
| I2 (p for heterogeneity) | 46.4% (0.001) | 53.3% (<0.0001) | 54.2% (0.002) | ||||||
| Recessive model | |||||||||
| Random | 1.34 | 1.11, 1.62 | 1.20 | 0.95, 1.51 | 1.24 | 1.07, 1.47 | |||
| I2 (p for heterogeneity) | 58% (<0.0001) | 14.8% (0.238) | 40.2% (0.03) | ||||||
| Dominant model | |||||||||
| Random | 1.15 | 1.04, 1.27 | 1.12 | 1.00, 1.26 | 1.17 | 1.05, 1.32 | |||
| I2 (p for heterogeneity) | 59.5% (<0.0001) | 55.8% (<0.0001) | 60.2% (<0.0001) | ||||||
The rare allele is Asp and the common is Glu.
The rare allele is a and the common is b.
The rare allele is C and the common is T.
CI, confidence interval.
Comparison | Glu298Asp | Intron 4 | -786T>C | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Odds ratio | 95% CI§ | Odds ratio | 95% CI | Odds ratio | 95% CI | ||||
| Additive model | |||||||||
| Random | 1.17 | 1.07, 1.28 | 1.12 | 1.01, 1.24 | 1.17 | 1.07, 1.28 | |||
| I2 (p for heterogeneity) | 67.9% (<0.0001) | 55.4% (<0.0001) | 62.7% (<0.0001) | ||||||
| Homozygous for rare allele vs. homozygous for common allele | |||||||||
| Random | 1.36 | 1.12, 1.66 | 1.24 | 0.97, 1.60 | 1.35 | 1.11, 1.64 | |||
| I2 (p for heterogeneity) | 60.1% (<0.0001) | 21.5% (0.148) | 52.6% (0.003) | ||||||
| Heterozygous vs. homozygous for common allele | |||||||||
| Random | 1.07 | 0.98, 1.17 | 1.10 | 0.98, 1.24 | 1.13 | 1.00, 1.26 | |||
| I2 (p for heterogeneity) | 46.4% (0.001) | 53.3% (<0.0001) | 54.2% (0.002) | ||||||
| Recessive model | |||||||||
| Random | 1.34 | 1.11, 1.62 | 1.20 | 0.95, 1.51 | 1.24 | 1.07, 1.47 | |||
| I2 (p for heterogeneity) | 58% (<0.0001) | 14.8% (0.238) | 40.2% (0.03) | ||||||
| Dominant model | |||||||||
| Random | 1.15 | 1.04, 1.27 | 1.12 | 1.00, 1.26 | 1.17 | 1.05, 1.32 | |||
| I2 (p for heterogeneity) | 59.5% (<0.0001) | 55.8% (<0.0001) | 60.2% (<0.0001) | ||||||
Comparison | Glu298Asp | Intron 4 | -786T>C | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Odds ratio | 95% CI§ | Odds ratio | 95% CI | Odds ratio | 95% CI | ||||
| Additive model | |||||||||
| Random | 1.17 | 1.07, 1.28 | 1.12 | 1.01, 1.24 | 1.17 | 1.07, 1.28 | |||
| I2 (p for heterogeneity) | 67.9% (<0.0001) | 55.4% (<0.0001) | 62.7% (<0.0001) | ||||||
| Homozygous for rare allele vs. homozygous for common allele | |||||||||
| Random | 1.36 | 1.12, 1.66 | 1.24 | 0.97, 1.60 | 1.35 | 1.11, 1.64 | |||
| I2 (p for heterogeneity) | 60.1% (<0.0001) | 21.5% (0.148) | 52.6% (0.003) | ||||||
| Heterozygous vs. homozygous for common allele | |||||||||
| Random | 1.07 | 0.98, 1.17 | 1.10 | 0.98, 1.24 | 1.13 | 1.00, 1.26 | |||
| I2 (p for heterogeneity) | 46.4% (0.001) | 53.3% (<0.0001) | 54.2% (0.002) | ||||||
| Recessive model | |||||||||
| Random | 1.34 | 1.11, 1.62 | 1.20 | 0.95, 1.51 | 1.24 | 1.07, 1.47 | |||
| I2 (p for heterogeneity) | 58% (<0.0001) | 14.8% (0.238) | 40.2% (0.03) | ||||||
| Dominant model | |||||||||
| Random | 1.15 | 1.04, 1.27 | 1.12 | 1.00, 1.26 | 1.17 | 1.05, 1.32 | |||
| I2 (p for heterogeneity) | 59.5% (<0.0001) | 55.8% (<0.0001) | 60.2% (<0.0001) | ||||||
The rare allele is Asp and the common is Glu.
The rare allele is a and the common is b.
The rare allele is C and the common is T.
CI, confidence interval.
Intron 4 polymorphism.
Thirty-one studies (31 articles: 9,925 cases and 9,407 controls) that evaluated the association between the intron 4 polymorphism and CHD were included in the meta-analysis (15, 22, 64, 65, 68, 72, 74–83, 92, 94–96, 98, 100, 101, 104–106, 108–110, 113, 114). The per-allele odds ratio for the intron 4 variant was 1.12 (95 percent CI: 1.01, 1.24; p = 0.02; Web appendix figure 4). There was evidence of heterogeneity between studies (I2 = 55.4 percent,
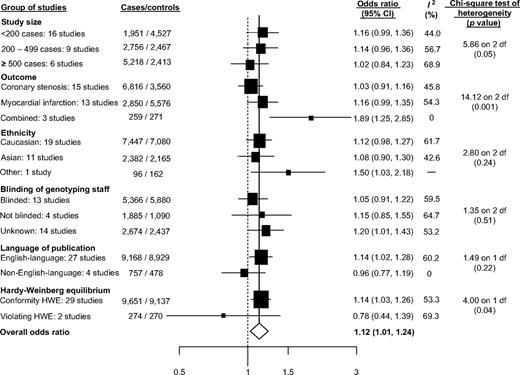
Studies of the intron 4 a/b variant and risk of coronary heart disease grouped by study characteristics. The sizes of the boxes relate to the inverse of the variance and thus to study size. CI, confidence interval; HWE, Hardy-Weinberg equilibrium.
-786T>C polymorphism.
Twenty-two studies (20 articles) that evaluated the association between the -786T>C polymorphism in the gene promoter and CHD were included in the meta-analysis (11,236 cases and 13,562 controls) (13, 22, 65, 66, 73, 79, 84–86, 89–92, 94, 107, 111, 113, 114, 117, 119). The per-allele odds ratio of CHD for the -786T>C variant was 1.17 (95 percent CI: 1.07, 1.28; p = 0.001; Web appendix figure 6). Substantial interstudy heterogeneity was observed (I2 = 62.7 percent,
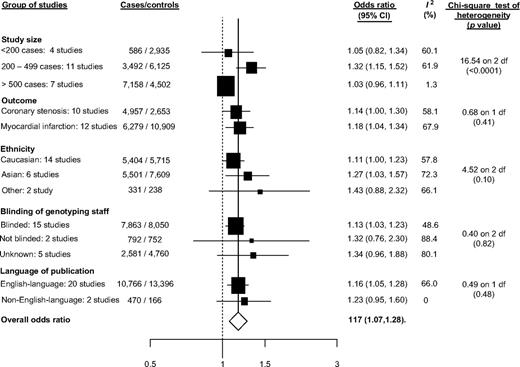
Studies of the -786T>C polymorphism and risk of coronary heart disease grouped by study characteristics. The sizes of the boxes relate to the inverse of the variance and thus to study size. CI, confidence interval.
Summary of the association between CHD and the NOS3 gene.
In these updated meta-analyses, several strategies were used to obtain unpublished data to minimize reporting and publication bias. An additional 28 studies and 7,840 cases for Glu298Asp, 15 studies and 3,713 cases for intron 4, and 14 studies and 8,859 cases for -786T>C were included in comparison with our previous report (17). Contrary to previous findings, in this updated meta-analysis, we observed statistical evidence of small-study bias in studies of the Glu298Asp and -786T>C polymorphisms. A stratified analysis of the associations of these two gene variants with CHD according to the number of cases supported these statistical findings and also suggested the presence of small-study bias for the intron 4 variant (figures 2 and 3). Interestingly, for Glu298Asp and intron 4, substantial heterogeneity was observed even within the group of studies with more than 500 cases (Glu298Asp: I2 = 61.4 percent; intron 4: I2 = 68.9 percent). In this updated meta-analysis, the -786T>C polymorphism was associated with an increased risk of CHD. For all three polymorphism-CHD associations, we observed substantial heterogeneity not explained by any of the study characteristics evaluated (figures 1, 2, and 3). Despite previous claims of a differential effect of ethnicity on gene-disease associations in complex diseases, in these meta-analyses, the mean estimate of the effects was highly consistent among the ethnic groups evaluated, a finding consistent with other recent results (129). A cumulative synthesis of NOS3 polymorphisms and CHD revealed that, for Glu298Asp and intron 4, the initial positive associations gradually attenuated over time and became more stable, as data accrued. However, for the -786T>C-CHD association, some degree of instability over time was observed (Web appendix figures 8, 9, and 10). Although other possible sources of bias (e.g., survival, classification, and selection) and confounding (cardiovascular risk factors) are possible, they are unlikely to be present (130). However, to confirm or refute a role of the NOS3 gene in CHD, future large-scale studies using a map-based “candidate gene” approach by genotyping tSNPs are warranted.
High blood pressure
Two linkage studies, in Caucasians, analyzed the sharing of alleles at the highly polymorphic dinucleotide repeat element in intron 13 of the NOS3 gene among hypertensive sibling pairs. Both studies failed to detect an excess of allele sharing that would implicate this region regarding susceptibility to hypertension (131, 132). Findings from an additional association substudy (using SNPs in introns 18 and 23) were also negative (131). Larger genome-wide linkage studies that used markers in the 7q35-36 region (region of the NOS3 gene) have also failed to find evidence of linkage to hypertension in this region (133, 134). Genetic association studies examining Glu298Asp, -786T>C, or intron 4 polymorphisms have yielded conflicting conclusions. As of January 2004, 18 studies (15 articles) were detected (42, 119–132). For details of the studies included, refer to Web appendix table 6.
The Glu298Asp polymorphism was the variant evaluated most frequently, with 12 studies (10 articles; 3,950 cases and 5,538 controls) (42, 135–143). Overall, in three of 12 studies, positive associations were reported (Shoji et al. (142): OR = 1.8, 95 percent CI: 1.1, 3.0; Miyamoto et al. (Kyoto) (140): OR = 2.3, 95 percent CI: 1.4, 3.9; and Miyamoto et al. (Kumamoto) (140): OR = 2.4, 95 percent CI: 1.4, 4.0) under a dominant model of inheritance. However, conflicting results have been reported, even in a larger study whose subjects were of the same ethnic background (143). The second most described NOS3 variant was -786T>C (four studies; 2,183 cases and 3,619 controls) (42, 143–145). Only one study in Caucasians (144) has been associated with an increased risk of hypertension (adjusted OR = 2.16, 95 percent CI: 1.3, 3.7) for individuals homozygous for the -786C allele compared with TT-genotype subjects; however, in the much larger, community-based case-control study in Japanese individuals by Tsujita et al. (143), a null association was found. Likewise, for the intron 4 polymorphism, conflicting results have been reported in six individually underpowered studies (total of 960 cases and 1,301 controls) published until January 2004 (136, 140, 142, 146–148). Subsequently, a large, population-based study in Caucasians found that the Glu298Asp polymorphism was not associated with either prevalent hypertension or difference in systolic or diastolic blood pressure by genotype (149).
In addition to the well-recognized difficulties in reliably identifying the small expected genotypic effect in complex disorders, there are additional difficulties in assessing blood pressure or hypertension as outcomes. Most of the previous studies used certain threshold blood pressures (e.g., ≥140 mmHg systolic or ≥90 mmHg diastolic) to define hypertension, assuming the presence of a clear cutoff point in the blood pressure-CHD association. Nevertheless, it has become clear that the risk associated with blood pressure is continuous and graded over the whole range of usually encountered blood pressure levels (150), even in those individuals considered normotensive. Use of blood pressure as a continuous outcome together with a gene-based approach using tSNPs may therefore be a more informative approach for assessing the effect of the NOS3 variation on blood pressure.
Preeclampsia
It has been proposed that enhanced synthesis of nitric oxide is responsible in part for the adaptive change in maternal hemodynamics status observed in normal pregnancy (151). Hypertensive disorders of pregnancy are characterized by an inappropriately high vascular resistance that might arise from reduced nitric oxide bioavailability (152, 153). Preeclampsia has a familial component, and women with disorders associated with endothelial dysfunction (e.g., hypertension, diabetes) are at greater risk of developing preeclampsia (154). Several authors have examined the role of the NOS3 gene in the pathogenesis of preeclampsia. In a recent meta-analysis of genetic association studies (12 studies; 1,334 cases and 2,894 controls) published up to November 2005, including a new case-control study (155), we did not detect a significant increase in the risk of preeclampsia associated with the Glu298Asp polymorphism under an additive model (OR = 1.03, 95 percent CI: 0.79, 1.34). Similar results were observed for a recessive model (OR = 1.28, 95 percent CI: 0.76, 2.16) and a dominant model (OR = 1.12, 95 percent CI: 0.84, 1.49) (155). Regarding other NOS3 polymorphisms (intron 4 and -786T>C), only a few case-control studies in different ethnic groups have evaluated the role of these polymorphisms (19, 156–158), and no increase in preeclampsia risk was observed (19).
Stroke
In a comprehensive meta-analysis of studies of all candidate genes for ischemic stroke in Caucasians published to January 2003, individuals homozygous for the Asp298 allele (three studies; 1,086 cases and 1,089 controls), in comparison with Glu298 carriers, did not have an increased risk of ischemic stroke (OR = 0.98, 95 percent CI: 0.76, 1.26) (159). Since that analysis, two additional studies among Caucasians have been published (160, 161); in both, no significant associations between the Glu298Asp genotypes and stroke were observed. In one of the studies (161), only those subjects with cerebral small-vessel disease were evaluated, and, despite null associations for the NOS3 Glu298Asp, -786T>C, and intron 4 polymorphisms overall, for a subgroup of patients with lacunar infarction, a protective effect for the a allele of intron 4 was reported. However, the possibility of a false-positive result because of multiple comparisons has to be considered. In the non-Caucasian population, the studies have mainly focused on the role of the intron 4 polymorphism and have yielded contradictory results. In the Chinese population, Hou et al. (162) (364 cases and 516 controls) observed an increased risk of ischemic stroke (OR = 2.13, 95 percent CI: 1.98, 4.80) for carriers of the a allele after adjusting for potential confounders. On the other hand, two other studies, one in Japanese and the other mainly in Caucasians (240 cases and 1,604 controls), found no increase in risk of stroke for carriers of the a allele (163, 164). In Afro-Americans, a recent, small case-control study (110 cases and 206 controls) of young women reported an increased risk of stroke for the -786T>C variant (OR = 2.9, 95 percent CI: 1.3, 6.4) (165).
Detailed analysis of studies reporting carotid stenosis as an outcome (87, 140, 166–172) can be found in the online Appendix. In addition, detailed analysis of studies reporting coronary spasm (40, 173–178), restenosis (179–182), diabetes mellitus (166, 183–185), renal disease (28, 186–192), and rheumatologic disorders (193–195) can also be found there.
GENE-ENVIRONMENT INTERACTIONS
Several interactions between the NOS3 polymorphisms and environmental factors have been proposed. Smoking has been the main focus of attention, particularly in studies of the intron 4 and -786T>C polymorphisms in Asian populations. Wang et al. (15) reported that smoking was an effect modifier of the intron 4–coronary stenosis association. Likewise, Nasreen et al. (196) found that, among smokers, homozygosity for the -786C allele was associated with a lower cerebral blood flow as well as higher cerebral vascular resistance in comparison with -786T allele carriers. Flow-mediated dilation was compared between healthy individuals in relation to the NOS3 Glu298Asp polymorphism by Leeson et al. (30), and interactions with a proatherogenic risk factor (smoking) and an antiatherogenic factor (n-3 fatty acids) were investigated. Flow-mediated dilation was not related to genotype in the group as a whole or within sexes. However, among men, smoking was associated with lower flow-mediated dilation in Asp298 carriers but not in Glu298 homozygotes. In the whole group, n-3 fatty acid levels were positively related to flow-mediated dilation in Asp298 carriers but not in Glu298 homozygotes. Thus, the Glu298Asp polymorphism may be associated with differences in the response of the endothelium to both smoking and n-3 fatty acid status. These early findings suggestive of gene-environmental interactions with the different NOS3 polymorphisms should be interpreted extremely cautiously, however, and much larger and more detailed studies are needed to clarify such putative interactions with appropriate power and rigor.
LABORATORY TESTS
All genetic association studies described in this review used genomic DNA extracted from blood. All involve amplification of genomic sequences containing the polymorphic sites. For determining the NOS3 Glu298Asp and -786T>C polymorphisms, the most frequently used method has been polymerase chain reaction and restriction fragment length polymorphism analysis (3). Other techniques have been a fluorescence or colorimetry-based allele-specific DNA-probe assay system (TaqMan; Applied Biosystems, Foster City, California) (12, 107). For the intron 4 polymorphism, the procedure used has been polymerase chain reaction. In most of the studies, quality control by DNA sequencing has been carried out in a small, random sample of the subjects genotyped in each study. Alternative methods for genotyping using melting curve analysis, in which the distinction of the different genotypes (wild type, mutant, and heterozygous) can be ascertained graphically by differences in their respective melting temperatures, have also been used (42).
POPULATION TESTING
We could not identify any published data on population testing of the NOS3 polymorphisms described in this review in relation to any of the associations described so far. The odds ratios for the Asp298 and 786C alleles are consistent with the genetic contribution to CHD being through small-to-moderate effects of many genes. Therefore, it seems unlikely that these polymorphisms individually will make a useful contribution to risk prediction in asymptomatic persons (17). However, whether combined genotype analysis integrated with orthodox assessment of cardiovascular risk will enhance the prediction of CHD warrants further exploration (197, 198).
CONCLUSIONS AND RECOMMENDATIONS FOR FUTURE RESEARCH
A considerable amount of evidence has accumulated evaluating the role of the NOS3 gene not only in cardiovascular disease but also in other complex disorders. An obstacle to evaluating, at the human level, the effects of the different NOS3 polymorphisms has been the lack of a reliable marker of the NOS3 gene function, partly due to the technical complexity in measuring nitric oxide production in humans. However, other than for CHD, no disease associations have been extensively described so far. Even for the positive associations with CHD, the likelihood of publication bias is substantial and cannot be excluded as a possible explanation. The complexity of the results observed for the association of each polymorphism with CHD, together with the absence of a definitive functional gene variant, strongly suggests that future studies evaluating the effect of the NOS3 gene on cardiovascular disease should use a “gene-based” approach. It seems unlikely that studies evaluating isolated NOS3 gene variants will produce a real advance in the understanding of the NOS3 gene and the role of nitric oxide. In addition, further and larger genetic association studies are needed in ethnic minorities with different allele/genotype frequencies of the NOS3 variants and different patterns of cardiovascular disease.
Editor's note:This paper is also available on the website of the Human Genome Epidemiology Network (http://www.cdc.gov/genomics/hugenet/).
A. D. H. holds a British Heart Foundation Senior Fellowship. S. H. holds the British Heart Foundation Chair of Cardiovascular Genetics (RG 2000/015). L. S. holds a Medical Research Council Clinician Scientist Fellowship. G. L. C. is supported by the Annals of Human Genetics Scholarship in human population genetics.
The authors thank Drs. X. L. Wang, M. Letonja, Y. Shimasaki, M. Nakayama, M. Yoshimura, H. Yasue, L. P. Lai, T. Wascher, A. Pulkkinen, J. Chen, D. Gu, A. Milutinovic, T. Ishigami, C. Zoccali, G. Tripepi, V. Dosenko, B. A. Nassar, Y. M. Lutay, M. H. Hutz, I. Jo, L. Afrasyap, D. Tousoulis, M. Kerkeni, G. P. Rossi, O. E. Mustafina, A. Gardemann, Y. Yamada, M. Tobin, M. Yokota, H. S. Kim, W. K. Park, E. Coto, V. Nicaud, F. Cambien, Y. L. Ko, C. Sekuri, F. S. Cam, A. Berdeli, C. Fatini, M. Yokota, Z. H. Yiyang, C. Mamotte, M. G. Colombo, L. A. Salazar-Navarrete, and C. Heltianu, who kindly provided additional valuable information from their studies to the meta-analyses. They also thank Carolina Paez for providing clerical support in obtaining relevant manuscripts.
A. D. H. is a co-inventor on a patent held by UCL for the use of ADMA (an inhibitor of nitric oxide synthase) as a diagnostic test for preeclampsia.
References
Stuehr DJ. Structure-function aspects in the nitric oxide synthases.
Marsden PA, Heng HH, Scherer SW, et al. Structure and chromosomal localization of the human constitutive endothelial nitric oxide synthase gene.
Hingorani AD. Polymorphisms in endothelial nitric oxide synthase and atherogenesis: John French Lecture 2000.
Mayer B, Hemmens B. Biosynthesis and action of nitric oxide in mammalian cells.
Charles IG, Scorer CA, Moro MA, et al. Expression of human nitric oxide synthase isozymes.
Vallance P, Leone A, Calver A, et al. Endogenous dimethylarginine as an inhibitor of nitric oxide synthesis.
Clarkson P, Celermajer DS, Powe AJ, et al. Endothelium-dependent dilatation is impaired in young healthy subjects with a family history of premature coronary disease.
Schachinger V, Britten MB, Elsner M, et al. A positive family history of premature coronary artery disease is associated with impaired endothelium-dependent coronary blood flow regulation.
Kuhlencordt PJ, Gyurko R, Han F, et al. Accelerated atherosclerosis, aortic aneurysm formation, and ischemic heart disease in apolipoprotein E/endothelial nitric oxide synthase double-knockout mice.
Hingorani AD, Liang CF, Fatibene J, et al. A common variant of the endothelial nitric oxide synthase (Glu298->Asp) is a major risk factor for coronary artery disease in the UK.
Nakayama M, Yasue H, Yoshimura M, et al. T(-786→C mutation in the 5′-flanking region of the endothelial nitric oxide synthase gene is associated with myocardial infarction, especially without coronary organic stenosis.
SeattleSNPs. Program for Genomic Applications. Supported by the National Heart, Lung, and Blood Institute, Seattle, WA. (URL: http://pga.gs.washington.edu). (Accessed January 1,
Wang XL, Sim AS, Badenhop RF, et al. A smoking-dependent risk of coronary artery disease associated with a polymorphism of the endothelial nitric oxide synthase gene.
Tanus-Santos JE, Desai M, Flockhart DA. Effects of ethnicity on the distribution of clinically relevant endothelial nitric oxide variants.
Casas JP, Bautista LE, Humphries SE, et al. Endothelial nitric oxide synthase genotype and ischaemic heart disease: meta-analysis of 26 studies involving 23028 subjects.
Rosas-Vargas H, Flores-Segura A, Guizada-Claure B, et al. Endothelial nitric oxide synthase gene polymorphism in the Indian and Mestizo populations of Mexico.
Serrano NC, Casas JP, Díaz LA, et al. eNOS genotyping and risk of preeclampsia: a multicentric case-control study.
Serrano NC, Diaz LA, Paez MC, et al. Endothelial nitric oxide synthase polymorphisms of clinical importance in cardiovascular illnesses: effect of ethnicity on the distribution in the Colombian population. (In Spanish). Bogotá, Colombia: COLCIENCIAS,
Jeerooburkhan N, Jones LC, Bujac S, et al. Genetic and environmental determinants of plasma nitrogen oxides and risk of ischemic heart disease.
International HapMap Project. (URL: http://www.hapmap.org/). (Accessed January 1,
Goldstein DB, Ahmadi KR, Weale ME, et al. Genome scans and candidate gene approaches in the study of common diseases and variable drug responses.
Weale ME, Depondt C, Macdonald SJ, et al. Selection and evaluation of tagging SNPs in the neuronal-sodium-channel gene SCN1A: implications for linkage-disequilibrium gene mapping.
Ahmadi KR, Weale ME, Xue ZY, et al. A single-nucleotide polymorphism tagging set for human drug metabolism and transport.
Tesauro M, Thompson WC, Rogliani P, et al. Intracellular processing of endothelial nitric oxide synthase isoforms associated with differences in severity of cardiopulmonary diseases: cleavage of proteins with aspartate vs. glutamate at position 298.
Persu A, Stoenoiu MS, Messiaen T, et al. Modifier effect of eNOS in autosomal dominant polycystic kidney disease.
Savvidou MD, Vallance PJ, Nicolaides KH, et al. Endothelial nitric oxide synthase gene polymorphism and maternal vascular adaptation to pregnancy.
Leeson CP, Hingorani AD, Mullen MJ, et al. Glu298Asp endothelial nitric oxide synthase gene polymorphism interacts with environmental and dietary factors to influence endothelial function.
Golser R, Gorren AC, Mayer B, et al. Functional characterization of Glu298Asp mutant human endothelial nitric oxide synthase purified from a yeast expression system.
McDonald DM, Alp NJ, Channon KM. Functional comparison of the endothelial nitric oxide synthase Glu298Asp polymorphic variants in human endothelial cells.
Fairchild TA, Fulton D, Fontana JT, et al. Acidic hydrolysis as a mechanism for the cleavage of the Glu(298)→Asp variant of human endothelial nitric-oxide synthase.
Rankinen T, Rice T, Perusse L, et al. NOS3 Glu298Asp genotype and blood pressure response to endurance training: the HERITAGE family study.
Naber CK, Baumgart D, Altmann C, et al. eNOS 894T allele and coronary blood flow at rest and during adenosine-induced hyperemia.
Philip I, Plantefeve G, Vuillaumier-Barrot S, et al. G894T polymorphism in the endothelial nitric oxide synthase gene is associated with an enhanced vascular responsiveness to phenylephrine.
Paradossi U, Ciofini E, Clerico A, et al. Endothelial function and carotid intima-media thickness in young healthy subjects among endothelial nitric oxide synthase Glu298→Asp and T-786→C polymorphisms.
Li R, Lyn D, Lapu-Bula R, et al. Relation of endothelial nitric oxide synthase gene to plasma nitric oxide level, endothelial function, and blood pressure in African Americans.
Miyamoto Y, Saito Y, Nakayama M, et al. Replication protein A1 reduces transcription of the endothelial nitric oxide synthase gene containing a -786T→C mutation associated with coronary spastic angina.
Nakayama M, Yasue H, Yoshimura M, et al. T-786→C mutation in the 5′-flanking region of the endothelial nitric oxide synthase gene is associated with coronary spasm.
Cattaruzza M, Guzik TJ, Slodowski W, et al. Shear stress insensitivity of endothelial nitric oxide synthase expression as a genetic risk factor for coronary heart disease.
Rossi GP, Taddei S, Virdis A, et al. The T-786C and Glu298Asp polymorphisms of the endothelial nitric oxide gene affect the forearm blood flow responses of Caucasian hypertensive patients.
Bilsborough W, Green DJ, Mamotte CD, et al. Endothelial nitric oxide synthase gene polymorphism, homocysteine, cholesterol and vascular endothelial function.
Tsukada T, Yokoyama K, Arai T, et al. Evidence of association of the ecNOS gene polymorphism with plasma NO metabolite levels in humans.
Wang XL, Sim AS, Wang MX, et al. Genotype dependent and cigarette specific effects on endothelial nitric oxide synthase gene expression and enzyme activity.
Yoon Y, Song J, Hong SH, et al. Plasma nitric oxide concentrations and nitric oxide synthase gene polymorphisms in coronary artery disease.
World Health Organization. The World Health Report
National Heart, Lung, and Blood Institute. Morbidity & mortality: 2002 chartbook on cardiovascular, lung and blood diseases. Rockville, MD: US Department of Health and Human Services, National Institutes of Health,
American Heart Association.
De Backer G, Ambrosioni E, Borch-Johnsen K, et al. European guidelines on cardiovascular disease prevention in clinical practice: third joint task force of European and other societies on cardiovascular disease prevention in clinical practice (constituted by representatives of eight societies and by invited experts).
Pearson TA, Blair SN, Daniels SR, et al. AHA Guidelines for Primary Prevention of Cardiovascular Disease and Stroke: 2002 Update: Consensus Panel Guide to Comprehensive Risk Reduction for Adult Patients Without Coronary or Other Atherosclerotic Vascular Diseases. American Heart Association Science Advisory and Coordinating Committee.
Goldstein LB, Adams R, Becker K, et al. Primary prevention of ischemic stroke: a statement for healthcare professionals from the Stroke Council of the American Heart Association.
Nieto FJ. Infections and atherosclerosis: new clues from an old hypothesis?
Homocysteine Studies Collaboration. Homocysteine and risk of ischemic heart disease and stroke: a meta-analysis.
Pearson TA, Mensah GA, Alexander RW, et al. Markers of inflammation and cardiovascular disease: application to clinical and public health practice: a statement for healthcare professionals from the Centers for Disease Control and Prevention and the American Heart Association.
Hajjar I, Kotchen TA. Trends in prevalence, awareness, treatment, and control of hypertension in the United States, 1988–2000.
Vasan RS, Larson MG, Leip EP, et al. Assessment of frequency of progression to hypertension in non-hypertensive participants in the Framingham Heart Study: a cohort study.
Kannel WB. Blood pressure as a cardiovascular risk factor: prevention and treatment.
Smith GC, Pell JP, Walsh D. Pregnancy complications and maternal risk of ischaemic heart disease: a retrospective cohort study of 129,290 births.
Colhoun HM, McKeigue PM, Davey Smith G. Problems of reporting genetic associations with complex outcomes.
Lohmueller KE, Pearce CL, Pike M, et al. Meta-analysis of genetic association studies supports a contribution of common variants to susceptibility to common disease.
Shadish WR, Haddock CK. Combining estimates of effect size. In: Cooper H, Hedges LV, eds. The handbook of research synthesis. New York, NY: Russell Sage Foundation,
Hibi K, Ishigami T, Tamura K. Endothelial nitric oxide synthase gene polymorphism and acute myocardial infarction.
Granath B, Taylor RR, Van Bockxmeer FM, et al. Lack of evidence for association between endothelial nitric oxide synthase gene polymorphisms and coronary artery disease in the Australian Caucasian population.
Shimasaki Y, Yasue H, Yoshimura M, et al. Association of the missense Glu298Asp variant of the endothelial nitric oxide synthase gene with myocardial infarction.
Wang CL, Hsu LA, Ko YS, et al. Lack of association between the Glu298Asp variant of the endothelial nitric oxide synthase gene and the risk of coronary artery disease among Taiwanese.
Yoon Y, Song J, Hong SH, et al. Plasma nitric oxide concentrations and nitric oxide synthase gene polymorphisms in coronary artery disease.
Aras O, Hanson NQ, Bakanay SM, et al. Endothelial nitric oxide gene polymorphism (Glu298Asp) is not associated with coronary artery disease in Turkish population.
Cai H, Wilcken DE, Wang XL. The Glu-298→Asp (894G→T) mutation at exon 7 of the endothelial nitric oxide synthase gene and coronary artery disease.
Cai H, Wang XL, Colagiuri S, et al. A common Glu298→Asp (894 G→T) mutation at exon 7 of the endothelial nitric oxide synthase gene and vascular complications in type 2 diabetes.
Gardemann A, Lohre J, Cayci S, et al. The T allele of the missense Glu(298)Asp endothelial nitric oxide synthase gene polymorphism is associated with coronary heart disease in younger individuals with high atherosclerotic risk profile.
Poirier O, Mao C, Mallet C, et al. Polymorphisms of the endothelial nitric oxide synthase gene—no consistent association with myocardial infarction in the ECTIM study.
Pulkkinen A, Viitanen L, Kareinen A, et al. Intron 4 polymorphism of the endothelial nitric oxide synthase gene is associated with elevated blood pressure in type 2 diabetic patients with coronary heart disease.
Ichihara S, Yamada Y, Fujimura T, et al. Association of a polymorphism of the endothelial constitutive nitric oxide synthase gene with myocardial infarction in the Japanese population.
Lee WH, Hwang TH, Oh GT, et al. Genetic factors associated with endothelial dysfunction affects the early onset of coronary artery disease in Korean males.
Nakagami H, Ikeda U, Maeda Y, et al. Coronary artery disease and endothelial nitric oxide synthase and angiotensin-converting enzyme gene polymorphisms.
Park JE, Lee WH, Hwang TH, et al. Aging affects the association between endothelial nitric oxide synthase gene polymorphism and acute myocardial infarction in the Korean male population.
Alvarez R, Gonzalez P, Batalla A, et al. Association between the NOS3 (-786 T/C) and the ACE (I/D) DNA genotypes and early coronary artery disease.
Cine N, Hatemi AC, Erginel-Unaltuna N. Association of a polymorphism of the ecNOS gene with myocardial infarction in a subgroup of Turkish MI patients.
Fowkes FG, Lee AJ, Hau CM, et al. Methylene tetrahydrofolate reductase (MTHFR) and nitric oxide synthase (ecNOS) genes and risks of peripheral arterial disease and coronary heart disease: Edinburgh Artery Study.
Hooper WC, Lally C, Austin H, et al. The relationship between polymorphisms in the endothelial cell nitric oxide synthase gene and the platelet GPIIIa gene with myocardial infarction and venous thromboembolism in African Americans.
Sigusch HH, Surber R, Lehmann MH, et al. Lack of association between 27-bp repeat polymorphism in intron 4 of the endothelial nitric oxide synthase gene and the risk of coronary artery disease.
Sim AS, Wang J, Wilcken D, et al. MspI polymorphism in the promoter of the human endothelial constitutive NO synthase gene in Australian Caucasian population.
Takagi S, Goto Y, Nonogi H, et al. Genetic polymorphisms of angiotensin converting enzyme (I/D) and endothelial nitric oxide synthase (T(-788)C) genes in Japanese patients with myocardial infarction.
Colombo MG, Paradossi U, Andreassi MG, et al. Endothelial nitric oxide synthase gene polymorphisms and risk of coronary artery disease.
Schmoelzer I, Renner W, Paulweber B, et al. Lack of association of the Glu298Asp polymorphism of endothelial nitric oxide synthase with manifest coronary artery disease, carotid atherosclerosis and forearm vascular reactivity in two Austrian populations.
Liyou N, Simons L, Friedlander Y, et al. Coronary artery disease is not associated with the E298→D variant of the constitutive, endothelial nitric oxide synthase gene.
Rossi GP, Cesari M, Zanchetta M, et al. The T-786C endothelial nitric oxide synthase genotype is a novel risk factor for coronary artery disease in Caucasian patients of the GENICA study.
Hirashiki A, Yamada Y, Murase Y, et al. Association of gene polymorphisms with coronary artery disease in low- or high-risk subjects defined by conventional risk factors.
Afrasyap L, Ozturk G. NO level and endothelial NO synthase gene polymorphism (Glu298Asp) in the patients with coronary artery disease from the Turkish population.
Agema WR, de Maat MP, Zwinderman AH, et al. An integrated evaluation of endothelial constitutive nitric oxide synthase polymorphisms and coronary artery disease in men.
Berdeli A, Sekuri C, Sirri Cam F, et al. Association between the eNOS (Glu298Asp) and the RAS genes polymorphisms and premature coronary artery disease in a Turkish population.
Fatini C, Sofi F, Sticchi E, et al. Influence of endothelial nitric oxide synthase gene polymorphisms (G894T, 4a4b, T-786C) and hyperhomocysteinemia on the predisposition to acute coronary syndromes.
Letonja M. The eNOS gene polymorphism does not have a major impact on lipid parameters and premature coronary artery disease in Caucasian women.
Park KW, You KH, Oh S, et al. Association of endothelial constitutive nitric oxide synthase gene polymorphism with acute coronary syndrome in Koreans.
Tobin MD, Braund PS, Burton PR, et al. Genotypes and haplotypes predisposing to myocardial infarction: a multilocus case-control study.
Yoon S, Shin C, Park HY, et al. Endothelial nitric oxide synthase gene is associated with vessel stenosis in Korean population.
Choi CJ, Lee KS, Baek SH, et al. Association of endothelial NO synthase gene Glu298Asp polymorphism with acute myocardial infarction.
Mustafina OE, Shagisultanova EI, Nasibullin TR, et al. Endothelial nitric oxide synthase gene minisatellite polymorphism: study in populations of the Volga-Ural region and analysis of associations with myocardial infarct and essential hypertension. (In Russian).
Spiridonova MG, Stepanov VA, Puzyrev VP, et al. Analysis of gene complexes predisposing to coronary atherosclerosis. (In Russian).
Sobstyl J, Dzida G, Puzniak A, et al. Analysis of association of human endothelial nitric oxide synthase gene polymorphism with myocardial infarction. (In Polish).
Wei D, Shan J, Chen Z, et al. The G894T mutation of the endothelial nitric oxide synthase gene is associated with coronary atherosclerotic heart disease in Chinese. (In Chinese).
Odawara M, Sasaki K, Tachi Y, et al. Endothelial nitric oxide synthase gene polymorphism and coronary heart disease in Japanese NIDDM.
Hwang JJ, Tsai CT, Yeh HM, et al. The 27-bp tandem repeat polymorphism in intron 4 of the endothelial nitric oxide synthase gene is not associated with coronary artery disease in a hospital-based Taiwanese population.
Wu YW, Lee CM, Hsu SM, et al. Association between endothelial nitric oxide synthase polymorphisms and the risk of premature coronary artery disease in Taiwan.
Yamada Y, Izawa H, Ichihara S, et al. Prediction of the risk of myocardial infarction from polymorphisms in candidate genes.
Heltianu C, Costache G, Gafencu A, et al. Relationship of eNOS gene variants to diseases that have in common an endothelial cell dysfunction.
Matyar S, Attila G, Acarturk E, et al. eNOS gene intron 4 a/b VNTR polymorphism is a risk factor for coronary artery disease in southern Turkey.
Milutinovic A, Hruskovicova H. The eNOS gene polymorphism does not have a major impact on lipid parameters and premature coronary artery disease in Slovene men (Caucasians).
Rios DL, Callegari-Jacques SM, Hutz MH. Endothelial nitric oxide synthase and fractalkine chemokine receptor polymorphisms on angiographically assessed coronary atherosclerosis.
Zhan YY, Di Q, Cheng YL, et al. Correlation between Glu298Asp polymorphism of vascular endothelial nitric oxide synthase gene and myocardial infarction in the elderly.
Lutay YM, Parkhomenko AN, Dovgan NV, et al. Endothelial nitric oxide synthase gene polymorphism in patients with acute coronary syndrome without ST segment elevation. (Abstract).
Dosenko VIe, Zahorii VIu, Lutai IaM, et al. Allelic polymorphism of endothelial NO-synthase (T(-786)→C) promoter gene as risk factor of acute coronary syndrome. (In Ukrainian).
Kerkeni M, Addad F, Chauffert M, et al. Hyperhomocysteinemia, endothelial nitric oxide synthase polymorphism, and risk of coronary artery disease.
Antoniades C, Tousoulis D, Vasiliadou C, et al. Genetic polymorphism on endothelial nitric oxide synthase affects endothelial activation and inflammatory response during the acute phase of myocardial infarction.
Nassar BA, Rockwood K, Kirkland SA, et al. Improved prediction of early-onset coronary artery disease using APOE epsilon4, BChE-K, PPARgamma2 Pro12 and ENOS T-786C in a polygenic model.
Testa A, Spoto B, Tripepi G, et al. The GLU298ASP variant of nitric oxide synthase interacts with asymmetric dimethyl arginine in determining cardiovascular mortality in patients with end-stage renal disease.
Chen J, Su S, Huang J, et al. Haplotype analysis of the endothelial nitric oxide synthase gene in relation to acute myocardial infarction.
Qi J, Dai C, Huang T, et al. A smoking-dependent risk of coronary artery disease associated with a polymorphism of the endothelial nitric oxide synthase gene.
Jaramillo PC, Muñoz MA, Lanas MC, et al. Endothelial nitric oxide synthase G894T gene polymorphism in Chilean subjects with coronary artery disease and controls.
Lee WH, Hwang TH, Park JE, et al. Analysis of the endothelial nitric oxide synthase and -fibrinogen gene polymorphism in the development of acute myocardial infarction in Korean men.
Colombo MG, Andreassi MG, Paradossi U, et al. Evidence for association of a common variant of the endothelial nitric oxide synthase gene (Glu298→Asp polymorphism) to the presence, extent, and severity of coronary artery disease.
Thomas S, Bruce C, Birkhead A, et al. Effect of ecNOS polymorphisms and coronary artery disease upon exhaled nitric oxide.
Cam SF, Sekuri C, Tengiz I, et al. The G894T polymorphism on endothelial nitric oxide synthase gene is associated with premature coronary artery disease in a Turkish population.
Asakimori Y, Yorioka N, Tanaka J, et al. Association between ENOS gene polymorphism and cardiovascular events in nondiabetic hemodialysis patients: a prospective study.
Sharan K, Surrey S, Ballas S, et al. Association of T-786C eNOS gene polymorphism with increased susceptibility to acute chest syndrome in females with sickle cell disease.
Choi SY, Yoo KH, Park JS, et al. eNOS gene polymorphism in patients with acute coronary syndrome or variant angina in Korean.
Ioannidis JP, Ntzani EE, Trikalinos TA. ‘Racial’ differences in genetic effects for complex diseases.
Casas JP, Bautista LE, Humphries SE, et al. Do meta-analyses of association studies of endothelial nitric oxide synthase (eNOS) variants and ischaemic heart disease provide conclusive answers? (Authors' reply).
Bonnardeaux A, Nadaud S, Charru A, et al. Lack of evidence for linkage of the endothelial cell nitric oxide synthase gene to essential hypertension.
Hunt SC, Williams CS, Sharma AM, et al. Lack of linkage between the endothelial nitric oxide synthase gene and hypertension.
Rice T, Rankinen T, Province MA, et al. Genome-wide linkage analysis of systolic and diastolic blood pressure: the Quebec Family Study.
Rice T, Rankinen T, Chagnon YC, et al. Genomewide linkage scan of resting blood pressure: HERITAGE Family Study. Health, risk factors, exercise training, and genetics.
Chen W, Srinivasan SR, Elkasabany A, et al. Combined effects of endothelial nitric oxide synthase gene polymorphism (G894T) and insulin resistance status on blood pressure and familial risk of hypertension in young adults: the Bogalusa Heart Study.
Benjafield AV, Morris BJ. Association analyses of endothelial nitric oxide synthase gene polymorphisms in essential hypertension.
Lacolley P, Gautier S, Poirier O, et al. Nitric oxide synthase gene polymorphisms, blood pressure and aortic stiffness in normotensive and hypertensive subjects.
Karvonen J, Kauma H, Kervinen K, et al. Endothelial nitric oxide synthase gene Glu298Asp polymorphism and blood pressure, left ventricular mass and carotid artery atherosclerosis in a population-based cohort.
Jachymova M, Horky K, Bultas J, et al. Association of the Glu298Asp polymorphism in the endothelial nitric oxide synthase gene with essential hypertension resistant to conventional therapy.
Miyamoto Y, Saito Y, Kajiyama N, et al. Endothelial nitric oxide synthase gene is positively associated with essential hypertension.
Kato N, Sugiyama T, Morita H, et al. Lack of evidence for association between the endothelial nitric oxide synthase gene and hypertension.
Shoji M, Tsutaya S, Saito R, et al. Positive association of endothelial nitric oxide synthase gene polymorphism with hypertension in northern Japan.
Tsujita Y, Baba S, Yamauchi R, et al. Association analyses between genetic polymorphisms of endothelial nitric oxide synthase gene and hypertension in Japanese: The Suita Study.
Hyndman ME, Parsons HG, Verma S, et al. The T-786→C mutation in endothelial nitric oxide synthase is associated with hypertension.
Kajiyama N, Saito Y, Miyamoto Y, et al. Lack of association between T-786→C mutation in the 5′-flanking region of the endothelial nitric oxide synthase gene and essential hypertension.
Rodriguez-Esparragon FJ, Rodriguez-Perez JC, Macias-Reyes A, et al. Peroxisome proliferator-activated receptor-gamma2-Pro12Ala and endothelial nitric oxide synthase–4a/bgene polymorphisms are associated with essential hypertension.
Uwabo J, Soma M, Nakayama T, et al. Association of a variable number of tandem repeats in the endothelial constitutive nitric oxide synthase gene with essential hypertension in Japanese.
Yokoyama K, Tsukada T, Nakayama M, et al. An intron 4 gene polymorphism in endothelial cell nitric oxide synthase might modulate volume-dependent hypertension in patients on hemodialysis.
Wolff B, Grabe HJ, Schluter C, et al. Endothelial nitric oxide synthase Glu298Asp gene polymorphism, blood pressure and hypertension in a general population sample.
Lewington S, Clarke R, Qizilbash N, et al. Prospective Studies Collaboration. Age-specific relevance of usual blood pressure to vascular mortality: a meta-analysis of individual data for one million adults in 61 prospective studies.
Williams DJ, Vallance PJ, Neild GH, et al. Nitric oxide-mediated vasodilation in human pregnancy.
Seligman SP, Buyon JP, Clancy RM, et al. The role of nitric oxide in the pathogenesis of preeclampsia.
Choi JW, Im MW, Pai SH. Nitric oxide production increases during normal pregnancy and decreases in preeclampsia.
Hingorani AD. Endothelial nitric oxide synthase polymorphisms and hypertension.
Yu CK, Casas JP, Savvidou MD, et al. Endothelial nitric oxide synthase gene polymorphism (Glu298Asp) and development of pre-eclampsia: a case-control study and a meta-analysis.
Bashford MT, Hefler LA, Vertrees TW, et al. Angiotensinogen and endothelial nitric oxide synthase gene polymorphisms among Hispanic patients with preeclampsia.
Tempfer CB, Dorman K, Deter RL, et al. An endothelial nitric oxide synthase gene polymorphism is associated with preeclampsia.
Grandone E, Colaizzo D, Martinelli P, et al. Does endothelial nitric oxide synthase gene variation play a role in the occurrence of hypertension in pregnancy?
Casas JP, Hingorani AD, Bautista LE, et al. Meta-analysis of genetic studies in ischaemic stroke: thirty-two genes involving approximately 18,000 cases and 58,000 controls.
Szolnoki Z, Havasi V, Bene J, et al. Endothelial nitric oxide synthase gene interactions and the risk of ischaemic stroke.
Hassan A, Gormley K, O'Sullivan M, et al. Endothelial nitric oxide gene haplotypes and risk of cerebral small-vessel disease.
Hou L, Osei-Hyiaman D, Yu H, et al. Association of a 27-bp repeat polymorphism in ecNOS gene with ischemic stroke in Chinese patients.
Yahashi Y, Kario K, Shimada K, et al. The 27-bp repeat polymorphism in intron 4 of the endothelial cell nitric oxide synthase gene and ischemic stroke in a Japanese population.
Austin H, Chimowitz MI, Hill HA, et al. Genetics and Stroke in the Young Study Group. Cryptogenic stroke in relation to genetic variation in clotting factors and other genetic polymorphisms among young men and women.
Howard TD, Giles WH, Xu J, et al. Promoter polymorphisms in the nitric oxide synthase 3 gene are associated with ischemic stroke susceptibility in young black women.
Ohtoshi K, Yamasaki Y, Gorogawa S, et al. Association of (-)786T-C mutation of endothelial nitric oxide synthase gene with insulin resistance.
Frost D, Chitu J, Meyer M, et al. Endothelial nitric oxide synthase (ecNOS) 4 a/b gene polymorphism and carotid artery intima-media thickness in type-1 diabetic patients.
Wolff B, Braun C, Schluter C, et al. Endothelial nitric oxide synthase Glu(298)→Asp polymorphism, carotid atherosclerosis and intima-media thickness in a general population sample.
Markus HS, Ruigrok Y, Ali N, et al. Endothelial nitric oxide synthase exon 7 polymorphism, ischemic cerebrovascular disease, and carotid atheroma.
Asakimori Y, Yorioka N, Tanaka J, et al. Effect of polymorphism of the endothelial nitric oxide synthase and apolipoprotein E genes on carotid atherosclerosis in hemodialysis patients.
Ghilardi G, Biondi ML, DeMonti M, et al. Independent risk factor for moderate to severe internal carotid artery stenosis: T786C mutation of the endothelial nitric oxide synthase gene.
Lembo G, De Luca N, Battagli C, et al. A common variant of endothelial nitric oxide synthase (Glu298Asp) is an independent risk factor for carotid atherosclerosis.
Bertrand ME, LaBlanche JM, Tilmant PY, et al. Frequency of provoked coronary arterial spasm in 1089 consecutive patients undergoing coronary arteriography.
Yoshimura M, Yasue H, Nakayama M, et al. Genetic risk factors for coronary artery spasm: significance of endothelial nitric oxide synthase gene T-786→C and missense Glu298Asp variants.
Yoshimura M, Yasue H, Nakayama M, et al. A missense Glu298Asp variant in the endothelial nitric oxide synthase gene is associated with coronary spasm in the Japanese.
Nakayama M, Yoshimura M, Sakamoto T, et al. Synergistic interaction of T-786→C polymorphism in the endothelial nitric oxide synthase gene and smoking for an enhanced risk for coronary spasm.
Chang K, Baek SH, Seung KB, et al. The Glu298Asp polymorphism in the endothelial nitric oxide synthase gene is strongly associated with coronary spasm.
Gomma AH, Elrayess MA, Knight CJ, et al. The endothelial nitric oxide synthase (Glu298Asp and -786T>C) gene polymorphisms are associated with coronary in-stent restenosis.
Gorchakova O, Koch W, von Beckerath N, et al. Association of a genetic variant of endothelial nitric oxide synthase with the 1 year clinical outcome after coronary stent placement.
Volzke H, Grimm R, Robinson DM, et al. Candidate genetic markers and the risk of restenosis after coronary angioplasty.
Suzuki T, Okumura K, Sone T, et al. The Glu298Asp polymorphism in endothelial nitric oxide synthase gene is associated with coronary in-stent restenosis.
Ukkola O, Erkkila PH, Savolainen MJ, et al. Lack of association between polymorphisms of catalase, copper-zinc superoxide dismutase (SOD), extracellular SOD and endothelial nitric oxide synthase genes and macroangiopathy in patients with type 2 diabetes mellitus.
Monti LD, Barlassina C, Citterio L, et al. Endothelial nitric oxide synthase polymorphisms are associated with type 2 diabetes and the insulin resistance syndrome.
Lee YJ, Chang DM, Tsai JC. Association of a 27-bp repeat polymorphism in intron 4 of endothelial constitutive nitric oxide synthase gene with serum uric acid levels in Chinese subjects with type 2 diabetes.
Zanchi A, Moczulski DK, Hanna LS, et al. Risk of advanced diabetic nephropathy in type 1 diabetes is associated with endothelial nitric oxide synthase gene polymorphism.
Shimizu T, Onuma T, Kawamori R, et al. Endothelial nitric oxide synthase gene and the development of diabetic nephropathy.
Rippin JD, Patel A, Belyaev ND, et al. Nitric oxide synthase gene polymorphisms and diabetic nephropathy.
Noiri E, Satoh H, Taguchi J, et al. Association of eNOS Glu298Asp polymorphism with end-stage renal disease.
Merta M, Reiterova J, Tesar V, et al. Influence of the endothelial nitric oxide synthase polymorphism on the progression of autosomal dominant polycystic kidney disease and IgA nephropathy.
Walker D, Consugar M, Slezak J, et al. The ENOS polymorphism is not associated with severity of renal disease in polycystic kidney disease 1.
Neugebauer S, Baba T, Watanabe T. Association of the nitric oxide synthase gene polymorphism with an increased risk for progression to diabetic nephropathy in type 2 diabetes.
Kim JU, Chang HK, Lee SS, et al. Endothelial nitric oxide synthase gene polymorphisms in Behcet's disease and rheumatic diseases with vasculitis.
Salvarani C, Boiardi L, Casali B, et al. Endothelial nitric oxide synthase gene polymorphisms in Behcet's disease.
Serrano NC, Paez C, Correa PA, et al. Endothelial nitric oxide synthase gene polymorphism is associated with systemic lupus erythematosus.
Nasreen S, Nabika T, Shibata H, et al. T-786C polymorphism in endothelial NO synthase gene affects cerebral circulation in smokers: possible gene-environmental interaction.
Yang Q, Khoury MJ, Botto L, et al. Improving the prediction of complex diseases by testing for multiple disease-susceptibility genes.


{kind=link}
{kind=link}
{kind=link}