
Abstract
Autism is a prevailing neurodevelopmental disorder with a large genetic/genomic component. Recently, the dual-specificity tyrosine-(Y)-phosphorylation-regulated kinase 1 A (DYRK1A) gene was implicated as a risk factor for autism spectrum disorder (ASD). We identified five DYRK1A variants in ASD patients and found that the dose of DYRK1A protein has a crucial role in various aspects of postnatal neural development. Dyrk1a loss of function and gain of function led to defects in dendritic growth, dendritic spine development and radial migration during cortical development. Importantly, two autism-associated truncations, R205X and E239X, were shown to be Dyrk1a loss-of-function mutants. Studies of the truncated Dyrk1a mutants may provide new insights into the role of Dyrk1a in brain development, as well as the role of Dyrk1a loss of function in the pathophysiology of autism.
Similar content being viewed by others
Introduction
Autism spectrum disorder (ASD) is a complex, heterogeneous neurodevelopmental disorder1 with multiple different etiologies. The heritability of ASD has been estimated to be as high as 90%.2 ASD is usually diagnosed by clinical assessment and the onset of three core disturbances before 3 years of age: impaired social interaction, unusual patterns of highly restricted interests and repetitive behaviors, and language abnormalities.2 The risk of recurrence for ASD is approximately 2–3%.3
Autism is thought to occur when an array of potential genetic vulnerabilities, possibly in concert with epigenetic factors and/or gene–environment interactions, affect neural connectivity and neurodevelopment.4 To date, we have learned that the symptoms of autism may arise when some specific region of the genome appears to be abnormal; for instance, multiple genetic/genomic factors have been reported to be associated with autism, including chromosomal abnormalities, genomic copy number variants and genetic variants of specific genes.5, 6, 7, 8
Dual-specificity tyrosine-(Y)-phosphorylation-regulated kinase 1A (DYRK1A), which is encoded by a gene on human chromosome 21, is involved in many of the biological processes of neural development and performs critical functions by phosphorylating various downstream substrates, including transcription factors, splice factors and synaptic proteins.9 For example, DYRK1A-mediated phosphorylation of the P53 protein at Ser15 inhibits cell proliferation and is therefore associated with cell growth and differentiation.10 The tau protein and the N-WASP protein affect the outgrowth of microscopic fibers and actin assembly when they are phosphorylated by DYRK1A and thereby contribute to the development of neuronal dendrites and dendritic spines.11, 12, 13 Considering the critical contribution of DYRK1A to Down syndrome (DS),14 numerous studies have focused on the Dyrk1a gain-of-function model, in which transgenic mice that overexpress Dyrk1a by 1.5-fold show severe learning and memory defects.15, 16, 17 The dosage imbalance of Dyrk1a led to delayed neural development.18, 19, 20 Previous studies have identified the molecular mechanisms by which DYRK1A affects the occurrence of DS and mental retardation.9, 15, 21, 22, 23 In several recent exome sequencing studies of ASD patients, some DYRK1A variants were found to be associated with autism, suggesting that DYRK1A is another risk factor for ASD.6, 24, 25, 26
In an exome sequencing pilot study, we identified several DYRK1A mutations in ASD patients with intellectual disability (ID) and developmental delay (DD) (short name ASD/ID/DD or ASD), consistent with the findings of other independent studies.27 Furthermore, we screened 892 patients with ASD/ID/DD and/or epilepsy/seizure for the DYRK1A gene and found additional variants of DYRK1A (Supplementary Table S1). Using this sequencing approach, a total of 11 novel variants and 1 known mutation in the coding region were detected, of which 9 variants were predicted to alter protein function according to bioinformatics prediction (Table 1), whereas the other 3 variants were benign. Given the locations of the DYRK1A variants in the specific protein domains, we postulate that those variants may affect DYRK1A function in three ways: nuclear localization, kinase activity, and abnormal translation termination that results in truncated DYRK1A proteins (Table 1 and Figure 1c). Previous reports have suggested that the R205X mutant acts in the third way.28, 29 Owing to the critical role of Dyrk1a in neural development, we decided to investigate the role of Dyrk1a in neural development, as well as whether these Dyrk1a variants have loss- or gain-of-function effects.

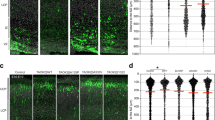
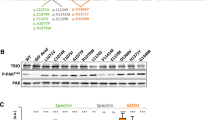
Wild-type (WT) Dyrk1a (dual-specificity tyrosine-(Y)-phosphorylation-regulated kinase 1 A) overexpression reduced total neurite and axon length of neurons in primary cultured mouse cortical neurons. (a) The Dyrk1a expression pattern in the cerebral cortex during the embryonic and postnatal period. (b) Specific knockdown of endogenous Dyrk1a in the mouse cortical neurons. Immunoblotting of the knockdown using specific short hairpin RNA (shRNA) against endogenous DYRK1A protein. The protein marker ladders are shown on the right. (c) Representation of the specific locations of missense variants related to autism in the DYRK1A protein structure, with the two truncations highlighted in red. The five variants located above the protein structure were included in the additional functional study. The smaller orange bar shows the NLS (bipartite nuclear localization signal) motif, and the larger bar (blue) represents the protein kinase domain. The black thread shows the other amino acids making up DYRK1A. (d) The specific cellular morphology of neurons expressing the CAG vector (as a control), Dyrk1a RNAi, Dyrk1a WT overexpression, the two truncations (R205X and E239X) and the three missense mutations (H119Y, A195T and L259F). All neurons were co-labeled with 4,6-diamidino-2-phenylindole (DAPI; to identify nuclei), green fluorescent protein (GFP; to identify overall neuronal morphology) and SMI 312 (an axonal marker). Scale bar 50 μm. (e, f) The statistical results for the total neurites (e) and axon length (f) between specific cell types. The bar graph shows the mean value±s.e.m.; approximately 70 cells from three independent experiments were counted during the statistical analysis. Unpaired t-test statistical method was used. **P<0.01, ***P<0.001, ****P<0.0001.
Materials and methods
Selected DYRK1A variants for the neuronal function study
Previously, we collaboratively screened 892 patients at Boston Children’s Hospital (437 samples of the western population) and the Children’s Hospital of Fudan University (455 samples of Han Chinese) with ASD/ID/DD for DYRK1A. All samples were de-identified prior to collection under Institutional Review Board protocols for non-identifiable biological specimens, which were deemed ‘leftover’ from patient samples after clinical tests. Of these, we selected samples with an ASD/ID/DD and/or epilepsy/seizure diagnosis according to Diagnostic and Statistical Manual of Mental Disorders, Fourth Edition. This study was approved by each institute. The sequencing experiments were also performed at each institute. For the bioinformatics analysis of the DYRK1A variants identified in the screen, the longest of DYRK1A isoform was used as a reference sequence (NM_001396.3, NP_001387.2). Considering the high homology between human and mouse Dyrk1a, we introduced the mouse Dyrk1a cDNA (NM_007890.2, NP_031916.1) into our study, and the DYRK1A variants in the coding region that were identified in individuals with ASD corresponded to Dyrk1a (Supplementary Table S2). Variants were classified according to DYRK1A protein domains listed in UniProt (http://www.uniprot.org/).
Supplementary Table S1 lists the variants detected in these patients. Based on the classification of the 11 variants, we selected 5 representative variants (three missense: H119Y, A195T and L259F; and two nonsense: R205X and E239X), which covered all three kinds of variants described previously, to investigate the role of DYRK1A in neurodevelopment, as well as the effects of these variants on the DYRK1A protein (Figure 1c). Considering that DYRK1A is highly conserved between human and mouse (approximately 99% according to protein sequence alignment using the MegAlign software; Supplementary Figure S1), we used the cDNA of mouse Dyrk1a rather than of human DYRK1A. The five variants were located in the mouse Dyrk1a cDNA, which corresponded to DYRK1A (Supplementary Table S2).
Clinical information on patients harboring DYRK1A mutations
The patient with R205X was diagnosed with ID and global DD. She exhibited microcephaly, chronic static encephalopathy and aphasia, oral motor apraxia, seizure dysmorphism and anxiety. All test results for karyotyping, fragile X and chromosomal microarray analysis were negative. The patient with E239X was diagnosed with ASD and global DD. He had a history of autism, anxiety and developmental coordination disorder, as well as abnormal electroencephalography but no seizures. Tests results for karyotyping, fragile X and chromosomal microarray analysis were negative. The patient with H119Y was diagnosed with PDD-NOS (pervasive developmental disorder, not otherwise specified). She had a history of global DD that was most marked for speech delay, motor delay and motor coordination disorder, seizure and mild physical dysmorphology. Tests results for karyotyping, fragile X and chromosomal microarray analysis were negative. Magnetic resonance imaging and magnetic resonance spectroscopy were normal. The patient with A195T was diagnosed with ASD/ID. He had a history of infantile spasms and global DD, with particular impairments in language, social interaction and repetitive behaviors, mild ID, complex partial epilepsy with epileptic encephalopathy and anxiety. Tests results for karyotyping, fragile X and chromosomal microarray analysis were negative. The patient with L259F was diagnosed with PDD-NOS and then ASD/ID. He demonstrated extremely low cognitive abilities and adaptive functioning, consistent with the mild range of ID. He had anxiety, perseveration and additional upsets and aggressive behavior. Because the parents of the patients were not available for testing, no other genetic information was available.
Plasmids
The full-length Dyrk1a ORF was amplified from the total cDNA of mouse cortical neurons. Dyrk1a was then inserted into pRK5-Myc, which already contained a 1xMyc epitope tag (5′-EQKLISEEDL-3′) at the 5′ end. For immunofluorescence experiments, we cloned Myc-Dyrk1a into pCAGGS-IRES-green fluorescent protein (GFP). The mutants, including both missense mutations and the two truncations, were generated using KOD-Plus, and specific primers were designed (Supplementary Table S3) according to the manufacturer’s instructions (#132000; Toyobo, Osaka, Japan). The mouse Dyrk1a shRNA was directed against the following sequence, 5′-AGAAGACGATGCATACAGG-3′, which was introduced into the pSUPER vector.
Lentivirus was provided by BrainVTA (Wuhan, China).
Antibodies
The antibodies used in this study included the following: DYRK1A (no. 2771; Cell Signaling Technology, Danvers, MA, USA); GAPDH (no. ab8245; Abcam, Cambridge, UK); Myc epitope tag (no. M20002; Abmart, Berkeley Heights, NJ, USA); GFP (no. G10362; Invitrogen, Carlsbad, CA, USA), and SMI 312 (no. SMI-312R; COVANCE, Princeton, NJ, USA).
Cell culture
Embryonic day 15 (E15)–E16 mouse cortical neurons were cultured and transfected via electroporation with an Amaxa Nucleofector (Amaxa, Cologne, Germany) at 0 days in vitro (DIV) on cover glasses that had been coated with a poly-D-lysine solution (1 mg ml−1 in dd water) according to the manufacturer’s instructions. After DIV 3, the cells were fixed for further analysis.
For neurite and axon outgrowth rescue experiments, we co-transfected each group with two plasmids, using a 1:1 ratio of wild-type (WT) or mutant forms of Dyrk1a and short hairpin RNAs (shRNAs), via electroporation as described above. Cells were collected for immunofluorescence analysis after 72 h of in vitro culturing.
Immunofluorescence
Cells were washed with 1 × phosphate-buffered saline (PBS) for 5 min, fixed in 4% paraformaldehyde (PFA) for 20 min and blocked in 1 × PBS buffer with 3% bovine serum albumin and 0.1% Triton-X-100 for 60 min at room temperature (RT). The cells were incubated in primary antibodies overnight at 4 °C, washed three times in 1 × PBS and then incubated in secondary antibodies at RT for 90–120 min. The signals were observed via fluorescence microscopy.
Immunoblotting
For Dyrk1a-specific expression in the mouse cortex, cortical tissue homogenates were prepared from the mouse brains at different developmental stages, immersed in RIPA buffer, boiled in 1 × sodium dodecyl sulfate loading buffer and then resolved by 8% sodium dodecyl sulfate-polyacrylamide gel electrophoresis. All of the protein on the gel was transferred to polyvinylidene fluoride membranes (Amersham Biosciences, Uppsala, Sweden), and the membranes were blocked with 5% non-fat milk in Tris-buffered saline and Tween 20 (TBST) buffer for 1 h. The membranes were then incubated in primary antibodies overnight at 4 °C, washed three times in TBST and incubated in secondary antibodies for 1 h at RT. The signals were revealed by horseradish peroxidase reaction using SuperSignal Chemiluminescent Substrate (Thermo Fisher Scientific, Waltham, MA, USA).
Hippocampal slice culture
Postnatal day 5 (P5) Sprague Dawley rats were included in the spine density study. Hippocampal tissue slices (400 μm) were cultured on Millicell (0.4 μm; Millipore, Boston, MA, USA) in six-well plates with 1 ml of slice culture medium as described previously.30, 31 The cultured slices were incubated in 5% CO2 at 37 °C and transfected using a Helios Gene Gun (Bio-Rad, Hercules, CA, USA) at DIV 2. Bullets were prepared containing 50 μg of plasmid DNA coated on 25 mg of 1.0-μm gold particles (Bio-Rad). The medium was changed every 2 days. The slices were fixed at DIV 7 in ice-cold 4% PFA. GFP immunostaining was then conducted using a GFP antibody and an Alexa 488-conjugated secondary antibody (1:1000). DAPI (4,6-diamidino-2-phenylindole; 1:500) was also used to visualize cell nuclei.
In utero electroporation
Plasmids encoding WT Dyrk1a, R205X and the empty vector were prepared in a final volume of 15 μl (2 μg μl−1 plasmid DNA, 0.01% Fast Green and 0.5 μg μl−1 enhanced GFP) for each pregnant mouse. The plasmids were then injected into the ventricles of E14.5 mouse brains. Electroporation (50-ms square pulses at 1000-ms intervals) was performed using an Electro Square Porator (ECM 830, San Diego, CA, USA) at 30 V.
Mouse pups were harvested at P0, and whole brains were fixed in 4% PFA for 24 h and sequentially dehydrated in 15% and 30% sucrose solutions (in 1 × PBS) for >48 h. Longitudinal brain sections (30 μm) were obtained via frozen sectioning for later immunofluorescence experiments in which GFP immunostaining was performed using a GFP antibody and an Alexa 488-conjugated secondary antibody (1:1000) to recognize GFP-positive neurons.
Data analysis
Images were obtained using a Nikon E80i fluorescence microscope (Nikon, Tokyo, Japan) equipped with a × 20 objective (for neurite development, axonal polarity formation and subcellular localization in cultured mouse cortical neurons), a Nikon E80i equipped with a × 10 objective (for cortical migration and in utero electroporation (IUE)), a Nikon A1 confocal microscope equipped with a × 20 objective (for neurite outgrowth in rat hippocampal slice cultures and the IUE examples) and a Nikon A1 confocal microscope equipped with a × 60 oil immersion lens (for spine density).
To evaluate neurite length in cultured primary cortical neurons, GFP-positive cells were randomly selected from each condition, and the total neurite length was traced with the Fiji software (http://imagej.net/Fiji/Downloads). At least three independent experiments were performed, and approximately 70 neurons were analyzed.
To evaluate spine density in rat hippocampal slice cultures, images were obtained as a Z-stack of approximately 15–20 sections (0.2 μm per section). The average spine density of specific neurons was analyzed statistically and compared among the four treatments. The average spine density was calculated as the total number of spines per 10 μm of dendritic length of each neuron. Approximately 45 neurons from 3 independent experiments were evaluated. The specific dendritic length was traced with the Fiji software (http://imagej.net/Fiji/Downloads).
To analyze total neurite outgrowth in slice cultures, images were obtained as a Z-stack of approximately 10–15 sections (1 μm per section). The total neurite length of each neuron was then traced with the Fiji software, as described previously.
For IUE, GFP-positive cells were counted using the ImageJ software (https://imagej.nih.gov/ij/download.html). The ratio of GFP-positive cells in the cortical plate included the total number of GFP-positive cells in the intermediate zone, the subventricular zone and the ventricular zone and was analyzed for each condition.
All statistical analyses were conducted using unpaired t-tests, with P<0.05 considered significant.
Results
Dyrk1a expression profiling during brain development
Dyrk1a is widely expressed in the developing nervous system,10, 32 suggesting that it has a crucial role in brain development. To examine DYRK1A protein levels in the embryonic and postnatal cortex, we collected protein lysates from embryonic and early postnatal mouse brains and examined DYRK1A protein levels via western blotting. We found that Dyrk1a was highly expressed during the embryonic stage, from E16 to P0, and this high expression level persisted until P7 (Figure 1a). In contrast, DYRK1A protein expression gradually decreased from P7 and remained at a relatively low level thereafter (Figure 1a). This expression pattern suggests that Dyrk1a has a critical role in the developing brain. To study the role of Dyrk1a in neural development, we designed a series of shRNAs against the mouse Dyrk1a gene and identified one set of effective shRNAs that significantly decreased DYRK1A protein levels in mouse primary cortical neurons (Figure 1b).
The dosage of Dyrk1a is crucial for dendritic growth in cultured neurons
Dendrites have a vital role in the biological functions of neurons, such as the receipt of information from other neurons. Previous studies have observed dendritic abnormalities in the brains of DS patients; in these patients, the DYRK1A gene is overexpressed owing to the excess of chromosome 21-associated genes.15, 18 Furthermore, Lepagnol-Bestel et al.21 showed that overexpressing Dyrk1a in mouse cortical neurons significantly decreased neurite length. Thus we decided to investigate whether autism-associated Dyrk1a mutations are involved in neurite development.
During our analysis of DYRK1A variants, we classified the nine variants into three groups according to their specific localization in the functional domains of DYRK1A (Figure 1c), as described above, and decided to select five representative mutants (R205X, E239X, H119Y, A195T and L259F) for further analysis.
In this study, we transfected mouse primary cortical neurons with plasmids expressing the empty CAG vector (as a control), Dyrk1a shRNA, WT Dyrk1a and the five Dyrk1a mutants, and we measured total neuritic and axonal length after 72 h. First, we confirmed a previous finding that overexpression of WT Dyrk1a severely reduced the total neurite length in cultured mouse cortical neurons (Figures 1d–f). Importantly, when endogenous Dyrk1a was specifically knocked down by RNA interference (RNAi), neurite length was also significantly decreased compared with control neurons expressing GFP alone. This finding was similar to the observation that overexpressing WT Dyrk1a decreased neurite growth (Figures 1d–f). These data indicate that the precise dosage of DYRK1A protein is critical for proper neurite development.
We further examined whether mutations in Dyrk1a affected neurite growth. We found that overexpressing either of the two Dyrk1a truncation mutants (R205X and E239X) did not inhibit total neurite growth; neurite growth in cells that were transfected with these mutants resembled that of controls, indicating that these two mutants lacked the normal function of WT Dyrk1a (Figures 1d–e). However, all three Dyrk1a missense mutants (H119Y, A195T and L259F) retained the ability to repress neurite outgrowth when overexpressed (Figures 1d–e), similar to WT Dyrk1a, indicating that non-haploinsufficient mutations may affect different aspects of brain development.
We next investigated whether Dyrk1a might affect axonal growth in neurons. During the neurite development assay, we measured axon length via immunohistochemistry using an antibody against the axonal marker SIM 312 and found that axonal growth was indeed affected by the Dyrk1a dosage and autism-associated mutations (Figures 1d and f). From these results, we found that shRNA-mediated Dyrk1a knockdown, as well as the overexpression of the WT form and the three missense mutants, decreased axon length compared with the control. In contrast, R205X- and E239X-expressing neurons exhibited the same phenotype as CAG vector-expressing neurons (Figures 1d and f).
Because WT Dyrk1a overexpression led to a similar phenotype to that of shRNA-mediated Dyrk1a knockdown, we performed a rescue experiment to confirm the specificity of the shRNA-mediated Dyrk1a knockdown and acquire a better understanding of the critical impact of the Dyrk1a dosage on neurite and axonal development. WT Dyrk1a rescued the inhibition of both neurite and axon length caused by Dyrk1a RNAi (Figures 2a–c). Neurons co-expressing WT Dyrk1a and shRNA exhibited similar outgrowth as control neurons expressing GFP alone, confirming the specificity of the Dyrk1a shRNA in mouse cortical neurons.

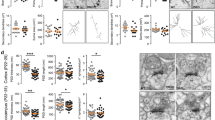
Wild-type (WT) Dyrk1a (dual-specificity tyrosine-(Y)-phosphorylation-regulated kinase 1 A) rescues the phenotype of decreased neurites and axon length in cultured mouse cortical neurons following Dyrk1a short hairpin RNA (shRNA) knockdown. (a) The specific morphology of neurons expressing the vectors (CAG and pSUPER), shRNA, WT Dyrk1a and WT or mutant forms of Dyrk1a along with the shRNA. All neurons were co-labeled with 4,6-diamidino-2-phenylindole (DAPI), green fluorescent protein (GFP) and SMI 312 (to distinguish axons from total neurites). Scale bar 50 μm. (b, c) The statistical results for the total neurites (b) and axon length (c) between specific cell types. The bar graph shows the mean value±s.e.m.; approximately 60 cells from three independent experiments were counted, and unpaired t-tests were used for the statistical analysis. ***P<0.001, ****P<0.0001.
Moreover, we previously showed that overexpression of nonsense mutations did not mimic the phenotype of WT Dyrk1a overexpression on neurite and axonal growth, revealing the important effect of these two Dyrk1a truncations. For further insight into the function of these mutants, we also conducted a rescue experiment using the five mutant forms of Dyrk1a and found that only the three missense mutations could rescue the knockdown phenotype (Figures 2a–c). The two truncations, R205X and E239X, did not restore neurite and axon lengths to control levels. In fact, staining revealed decreased outgrowth, similar to that observed with Dyrk1a RNAi (Figures 2a–c). Therefore, we surmise that these two Dyrk1a truncations act as Dyrk1a loss-of-function mutants.
We conclude that the Dyrk1a dosage is critical for proper neurite and axonal outgrowth and that the two nonsense mutations, R205X and E239X, are loss-of-function mutants.
Dyrk1a is crucial for the growth of neurites and spines in hippocampal neurons in slices
Next we wanted to examine the role of Dyrk1a in spine formation in hippocampal neurons. Dendritic spines are small protrusions along dendrites and represent the typical postsynaptic structure of excitatory synapses. Studies have shown that dendritic spine abnormalities are strongly associated with various neurological and neuropsychiatric disorders.33 Dyrk1a is reported to phosphorylate and inactivate the N-WASP protein, suggesting that Dyrk1a has a role in spine formation.12
We investigated the role of Dyrk1a in spine formation using a hippocampal slice culture system. We used gene-gun-mediated delivery to introduce plasmids encoding GFP along with WT Dyrk1a, Dyrk1a R205X and Dyrk1a shRNA into hippocampal neurons in postnatal hippocampal slices. Interestingly, we found that either knocking down Dyrk1a or overexpressing the WT form strongly repressed dendritic spine formation in hippocampal neurons, further indicating that a precise amount of DYRK1A protein is required for proper spine development (Figures 3a and b). However, overexpression of the R205X mutant did not similarly inhibit dendritic growth (Figures 3a and b).

Dyrk1a (dual-specificity tyrosine-(Y)-phosphorylation-regulated kinase 1 A) dosage imbalance reduced dendritic spine density in rat hippocampal slice culture, and wild-type (WT) Dyrk1a rescued the phenotype caused by Dyrk1a knockdown. (a, c) Morphology of the spine density under specific conditions with the expression of different plasmids, including CAG vector, Dyrk1a short hairpin RNA (shRNA), Dyrk1a WT, R205X (a) and shRNA rescue using WT or mutant Dyrk1a (c). Scale bar 5 μm. (b, d) Statistical analysis of the spine density (per 10 μm) in specific types of neurons. Forty-five neurons from three independent experiments were counted, and the individual neuronal spine density was determined. Unpaired t-tests were performed for the statistical analysis to compare each condition. **P<0.01, ***P<0.001, ****P<0.0001.
Furthermore, we conducted rescue assays to confirm the critical role of Dyrk1a in spine formation. WT Dyrk1a successfully rescued the loss of function, and neurons that received this treatment exhibited phenotypes similar to those of neurons that only expressed the CAG vector (Figures 3c and d). Therefore, we concluded that the shRNA against Dyrk1a was specific in rat hippocampal neuron. The three missense mutations also rescued the decreased spine density caused by Dyrk1a RNAi, whereas the two truncation mutants, R205X and E239X, did not rescue the effects on spine formation (Figures 3c and d), consistent with the rescue phenotype that was observed in mouse cortical neurons.
We also measured neurite outgrowth in hippocampal neurons that received the aforementioned manipulations. Consistently, either RNAi or WT Dyrk1a overexpression strongly inhibited neurite growth in hippocampal neurons in brain slices (Figures 4a and b). However, overexpression of the Dyrk1a R205X mutant did not affect neurite growth compared with WT Dyrk1a (Figures 4a and b). We next re-expressed WT and different Dyrk1a mutants along with RNAi in hippocampal slice cultures to assess whether these defects could be rescued. Only WT Dyrk1a and the missense mutants, but not R205X and E239X, were able to rescue the defects caused by RNAi (Figures 4c and d). The rescue results further confirmed the role of Dyrk1a in neurite development in hippocampal slices and that the truncations resulted in Dyrk1a loss of function. This evidence again indicates that the autism-related Dyrk1a truncations (R205X and E239X) are loss-of-function mutations.

Dyrk1a (dual-specificity tyrosine-(Y)-phosphorylation-regulated kinase 1 A) inhibited total neurite outgrowth in rat hippocampal neurons, and short hairpin RNA (shRNA)-mediated loss of function was rescued by wild-type Dyrk1a. (a, c) The morphology of neurons in each condition expressing the indicated plasmids. (a) Neurons expressed CAG vector, Dyrk1a short hairpin RNA (shRNA), Dyrk1a wild-type (WT) and R205X. (c) Neurons expressed specific plasmids for the rescue conditions, including WT and the five mutant forms of Dyrk1a plus Dyrk1a shRNA. (b, d) Statistical analysis of total neurite length in overexpression (b) and rescue conditions (d). Scale bar 50 μm. Approximately 50 neurons from three independent experiments were counted, and the total neurite length of each cell was determined. Unpaired t-tests were performed for the statistical analysis. **P<0.01, ***P<0.001, ****P<0.0001.
Dyrk1a is critical for cortical migration during early postnatal development
To further understand the role of Dyrk1a in embryonic and postnatal cortical development in the mouse brain, we examined cortical migration by manipulating Dyrk1a via IUE in mice.
Plasmids encoding GFP plus either the WT Dyrk1a or the R205X mutant were electroporated into the ventricular zone of E14.5 mouse brains, and the migration status of the GFP-labeled neurons was observed in neonatal pups on P0. Overexpression of WT Dyrk1a significantly delayed cortical migration compared with GFP expression alone (Figures 5a and b). In contrast, expressing the Dyrk1a R205X truncation in migrating neurons did not similarly affect cortical migration, again suggesting that the R205X truncation has a loss-of-function effect (Figures 5a and b). This evidence again indicates that the R205X truncation may be a loss-of-function mutant of Dyrk1a, which affects various aspects of postnatal neural development, including dendrite and spine formation and cortical migration.

Dyrk1a (dual-specificity tyrosine-(Y)-phosphorylation-regulated kinase 1 A) wild-type (WT) overexpression delayed neuronal migration in mouse embryonic cortical development. (a) Cortical migration in each condition, including CAG (left), Dyrk1a WT overexpression (middle) and R205X (right). (b) Quantification of panel (a). The green fluorescent protein (GFP)-positive cells in the cortical plate (CP), intermediate zone (IZ), subventricular zone (SVZ) and ventricular zone (VZ) were counted, and the ratio of GFP-positive cells in the CP/(IZ and VZ/SVZ) was analyzed statistically using unpaired t-tests. P0 mice were collected in three independent experiments, including CAG (n=7), Dyrk1A WT (n=6) and R205X (n=5) mice. ****P<0.0001. Scale bar 100 μm.
Autism-related Dyrk1a truncations do not affect subcellular localization
The histidine-repeat domain near the C-terminus of DYRK1A is critical for its localization in nuclear speckles.34 Therefore, we wanted to examine whether the R205X or E239X truncation affected the subcellular localization of Dyrk1a in neurons. We transfected cultured mouse primary cortical neurons with plasmids encoding myc-tagged WT Dyrk1a or the two truncated mutants. By performing immunocytochemistry with an anti-myc antibody, we first confirmed that WT Dyrk1a was expressed exclusively in the nucleus (Figure 6). Importantly, the R205X and E239X Dyrk1a truncations were also located in the nucleus, suggesting that these mutations did not affect Dyrk1a localization (Figure 6).

The two Dyrk1a (dual-specificity tyrosine-(Y)-phosphorylation-regulated kinase 1 A) truncations did not disturb Dyrk1a subcellular localization. (a–e) Neurons expressing Dyrk1a wild-type (WT) were labeled with 4,6-diamidino-2-phenylindole (DAPI; to identify nuclei), a Myc epitope tag (to identify the precise location of the expressed protein) and green fluorescent protein (GFP; to identify overall neuronal morphology). Merge 1 shows DAPI and Myc co-labeling, and merge 2 shows DAPI, Myc and GFP labeling. Panels (f–j) and (k–o) depict the two nonsense mutants, respectively. Both R205X and E239X, as well as Dyrk1a WT, localized to the nucleus. The images were merged using the ImageJ software. Scale bar 20 μm.
Discussion
DYRK1A is a critical gene located on human chromosome 21, encoding a proline-directed serine/threonine kinase that has been reported to phosphorylate several substrates related to gene transcription, neuronal degeneration and synapse function.9 Dyrk1a transgenic mice (TgDyrk1A), a mouse model of DS that overexpresses Dyrk1a by 1.5-fold,19 show defects in learning and memory, as well as hyperactivity and altered motor learning.22 Duplications of Dyrk1a result in abnormal neural development, such as compromised dendrite outgrowth and cell differentiation. At early embryonic stages, Dyrk1a overexpression reduces the proliferation of neuronal progenitor cells,35, 36, 37, 38, 39, 40 whereas increasing neuronal differentiation and enhancing GABAergic differentiation, which were observed in neuronal stem cells isolated from TgDyrk1A mice.
For decades, different types of genetic/genomic defects have been linked to ASD, including chromosomal abnormalities, genomic copy number variants and genetic variants of hundreds genes, highlighting the etiological heterogeneity underlying the genetic and phenotypic variation in ASD. Recently, DYRK1A was proposed as a candidate gene for syndromic ASD and ID or DD, and approximately 100 alterations have thus far been identified. Of these, the majority are de novo heterozygous mutations (nonsense, frame-shift, splicing alteration) and large chromosomal rearrangements (micro-deletion/duplication, insertion and translocation). The majority were haploinsufficiency alterations, which are relevant for many ASD-associated genes. The eight novel and one recurrent DYRK1A mutations detected in our patients with ASD/ID/DD are consistent with these findings.6, 8, 24, 25, 26, 41, 42, 43, 44
In the present study, the phenotypes of the two patients with DYRK1A truncations partially overlapped with the findings in a recent report, including microcephaly, seizures and impaired speech, supporting the hypothesis that DYRK1A truncations define a syndromic form of ASD and ID.26 Interestingly, the higher incidence of truncations identified in our patients (0.22%; 2 truncations in 892 patients) is similar to work by van Bon et al.26 (0.21%; 15 truncations in 7162 patients), further supporting the observation that DYRK1A truncations may be a relatively common type of mutation associated with syndromic ASD and ID or DD.
Fewer than 10 missense mutations have been reported in the literature, and most are predicted to be pathogenic; however, no functional studies have been described. In the present study, we found that the three missense mutations behaved similarly to WT Dyrk1a when overexpressed, with all three repressing neurite and axon outgrowth and affecting spine development; however, all could be rescued, which indicated that they were not loss-of-function mutations but may affect other Dyrk1a functions that were not assessed in this study. These findings also suggested that non-haploinsufficient missense mutations may affect brain development in different ways than haploinsufficient truncated mutations. Further studies to explore the pathogenesis of such missense mutations are needed. In addition, this study was limited to detecting only DYRK1A mutations; therefore, these missense mutations may in fact function with other unknown mutations or may not contribute to the risk of autism.
In this study, we show that Dyrk1a has an important role in neural development. The imbalance of the Dyrk1a dosage affected dendrite development and axon length. Consistent results were found in primary cultured mouse cortical neuron and rat hippocampal slice cultures. We also found that the two Dyrk1a nonsense truncations were unable to rescue defects caused by Dyrk1a knockdown. Therefore, we conclude that nonsense mutations caused a loss of function of the DYRK1A protein, which had effects on dendritic outgrowth in neurons, dendritic spine development and embryonic cortical migration. Moreover, upregulation and downregulation of Dyrk1a also reduced the dendritic spine density in rat hippocampal slice cultures. Importantly, cortical migration during the mouse embryonic stage was also altered by the overexpression of WT Dyrk1a from E14.5 to P0. Based on these results, we conclude that Dyrk1a has a key role in embryonic development.
Furthermore, overexpression of the two nonsense truncations resembled defects caused by Dyrk1a knockdown, suggesting that patients carrying Dyrk1a nonsense mutations show haploinsufficiency of Dyrk1a. Reduced DYRK1A proteins levels of in patients likely lead to defects in neural development and further ASD symptoms, providing new evidence that the dosage of Dyrk1a is critical for proper neurite and axonal outgrowth.
To date, the impact of Dyrk1a loss of function on neural development remains limited. For example, mice expressing only one copy of Dyrk1a exhibit a microcephalic phenotype.45, 46 However, the functional consequences of Dyrk1a loss of function have not been fully addressed. Dyrk1a knockout was reported to be lethal during the embryonic period, whereas Dyrk1a+/− mice survive but display severe defects in cognition, DDs and abnormalities in brain size and structure.47, 48 Thus it is difficult to undertake further analyses of this gene in a loss-of-function mouse model. The two Dyrk1a truncations we found in ASD patients provide important insights into the role of loss-of-function forms of Dyrk1a in various aspects of neural development.
DYRK1A phosphorylates multiple substrates. For instance, P53 Ser15 is directly phosphorylated by DYRK1A, and P53 phosphorylation at this site inhibits the proliferation of neural progenitors.10 DYRK1A has been shown to selectively phosphorylate RCAN1 at Ser112, prompting the phosphorylation of another residue, Ser108, by GSK3β and resulting in the formation of insoluble aggregates that contribute to the development of Alzheimer disease.11 N-WASP Thr192 may also be a substrate for DYRK1A phosphorylation, and spine formation can be inhibited by phosphorylation at this site.12 DYRK1A phosphorylates Tau at multiple sites, leading to decreased microtubule assembly and the formation of interneuron aggregates.13, 23 These observations reveal the potential role of Dyrk1a in dendritic outgrowth. In addition, DYRK1A negatively regulates nuclear factor of activated T cells by phosphorylating the serine-proline repeats 3 domain.49 Further studies are required to illustrate the exact substrates of DYRK1A that are responsible for various aspects of neural development.
In the present study, we showed that both excessive and reduced Dyrk1a activity yield similar phenotypes of delayed neural development that were distinct from the phenotypes associated with other candidate genes in neurodevelopment.50 Although ASD and DS are both neuronal development disorders and although previous studies have mostly focused on the relationship between DYRK1A and DS,9, 15, 21, 23 there are essential differences between ASD and DS. For example, the incidence stage and the specific phenotypes differ; severe mental retardation and delayed development is observed in DS patients, whereas verbal dysfunction and social problems are often observed in ASD patients.2 Genes in the DSCR region of chromosome 21 have clear connections with DS, whereas genetic factors other than chromosome 21 are associated with ASD. Thus it is important and necessary to identify the mechanisms by which DYRK1A contributes to ASD.
In summary, we found that Dyrk1a strongly affected various aspects of neuronal development, such as dendritic outgrowth, dendritic spine density and cortical migration, during early postnatal development. Importantly, R205X and E239X, two DYRK1A truncations that were identified in ASD/ID/DD patients, functioned as loss-of-function mutants.
References
Blenner S, Reddy A, Augustyn M . Diagnosis and management of autism in childhood. BMJ 2011; 343: d6238.
Zafeiriou DI, Ververi A, Dafoulis V, Kalyva E, Vargiami E . Autism spectrum disorders: the quest for genetic syndromes. Am J Med Genet B Neuropsychiatr Genet 2013; 162B: 327–366.
Geschwind DH, Levitt P . Autism spectrum disorders: developmental disconnection syndromes. Curr Opin Neurobiol 2007; 17: 103–111.
Geschwind DH . Autism: many genes, common pathways? Cell 2008; 135: 391–395.
Betancur C . Etiological heterogeneity in autism spectrum disorders: more than 100 genetic and genomic disorders and still counting. Brain Res 2011; 1380: 42–77.
O'Roak BJ, Vives L, Fu W, Egertson JD, Stanaway IB, Phelps IG et al. Multiplex targeted sequencing identifies recurrently mutated genes in autism spectrum disorders. Science 2012; 338: 1619–1622.
Dykens EM, Sutcliffe JS, Levitt P . Autism and 15q11-q13 disorders: behavioral, genetic, and pathophysiological issues. Ment Retard Dev Disabil Res Rev 2004; 10: 284–291.
Weiss LA, Shen Y, Korn JM, Arking DE, Miller DT, Fossdal R et al. Association between microdeletion and microduplication at 16p11.2 and autism. N Engl J Med 2008; 358: 667–675.
Park J, Song WJ, Chung KC . Function and regulation of Dyrk1A: towards understanding Down syndrome. Cell Mol Life Sci 2009; 66: 3235–3240.
Park J, Oh Y, Yoo L, Jung MS, Song WJ, Lee SH et al. Dyrk1A phosphorylates p53 and inhibits proliferation of embryonic neuronal cells. J Biol Chem 2010; 285: 31895–31906.
Song WJ, Song EA, Choi SH, Baik HH, Jin BK, Kim JH et al. Dyrk1A-mediated phosphorylation of RCAN1 promotes the formation of insoluble RCAN1 aggregates. Neurosci Lett 2013; 554: 135–140.
Park J, Sung JY, Park J, Song WJ, Chang S, Chung KC . Dyrk1A negatively regulates the actin cytoskeleton through threonine phosphorylation of N-WASP. J Cell Sci 2012; 125 (Pt 1): 67–80.
Liu F, Liang Z, Wegiel J, Hwang YW, Iqbal K, Grundke-Iqbal I et al. Overexpression of Dyrk1A contributes to neurofibrillary degeneration in Down syndrome. FASEB J 2008; 22: 3224–3233.
Ferrer I, Barrachina M, Puig B, Martinez de Lagran M, Marti E, Avila J et al. Constitutive Dyrk1A is abnormally expressed in Alzheimer disease, Down syndrome, Pick disease, and related transgenic models. Neurobiol Dis 2005; 20: 392–400.
Smith DJ, Stevens ME, Sudanagunta SP, Bronson RT, Makhinson M, Watabe AM et al. Functional screening of 2 Mb of human chromosome 21q22.2 in transgenic mice implicates minibrain in learning defects associated with Down syndrome. Nat Genet 1997; 16: 28–36.
Ahn KJ, Jeong HK, Choi HS, Ryoo SR, Kim YJ, Goo JS et al. DYRK1A BAC transgenic mice show altered synaptic plasticity with learning and memory defects. Neurobiol Dis 2006; 22: 463–472.
Altafaj X, Dierssen M, Baamonde C, Marti E, Visa J, Guimera J et al. Neurodevelopmental delay, motor abnormalities and cognitive deficits in transgenic mice overexpressing Dyrk1A (minibrain), a murine model of Down's syndrome. Hum Mol Genet 2001; 10: 1915–1923.
Scales TM, Lin S, Kraus M, Goold RG, Gordon-Weeks PR . Nonprimed and DYRK1A-primed GSK3 beta-phosphorylation sites on MAP1B regulate microtubule dynamics in growing axons. J Cell Sci 2009; 122 (Pt 14): 2424–2435.
Mazur-Kolecka B, Golabek A, Kida E, Rabe A, Hwang YW, Adayev T et al. Effect of DYRK1A activity inhibition on development of neuronal progenitors isolated from Ts65Dn mice. J Neurosci Res 2012; 90: 999–1010.
Kurabayashi N, Sanada K . Increased dosage of DYRK1A and DSCR1 delays neuronal differentiation in neocortical progenitor cells. Genes Dev 2013; 27: 2708–2721.
Lepagnol-Bestel AM, Zvara A, Maussion G, Quignon F, Ngimbous B, Ramoz N et al. DYRK1A interacts with the REST/NRSF-SWI/SNF chromatin remodelling complex to deregulate gene clusters involved in the neuronal phenotypic traits of Down syndrome. Hum Mol Genet 2009; 18: 1405–1414.
Arque G, Casanovas A, Dierssen M . Dyrk1A is dynamically expressed on subsets of motor neurons and in the neuromuscular junction: possible role in Down syndrome. PLoS One 2013; 8: e54285.
Ryoo SR, Jeong HK, Radnaabazar C, Yoo JJ, Cho HJ, Lee HW et al. DYRK1A-mediated hyperphosphorylation of Tau. A functional link between Down syndrome and Alzheimer disease. J Biol Chem 2007; 282: 34850–34857.
O'Roak BJ, Vives L, Girirajan S, Karakoc E, Krumm N, Coe BP et al. Sporadic autism exomes reveal a highly interconnected protein network of de novo mutations. Nature 2012; 485: 246–250.
Krumm N, O'Roak BJ, Shendure J, Eichler EE . A de novo convergence of autism genetics and molecular neuroscience. Trends Neurosci 2014; 37: 95–105.
van Bon BW, Coe BP, Bernier R, Green C, Gerdts J, Witherspoon K et al. Disruptive de novo mutations of DYRK1A lead to a syndromic form of autism and ID. Mol Psychiatry 2015; 21: 126–132.
De Rubeis S, He X, Goldberg AP, Poultney CS, Samocha K, Cicek AE et al. Synaptic, transcriptional and chromatin genes disrupted in autism. Nature 2014; 515: 209–215.
Redin C, Gerard B, Lauer J, Herenger Y, Muller J, Quartier A et al. Efficient strategy for the molecular diagnosis of intellectual disability using targeted high-throughput sequencing. J Med Genet 2014; 51: 724–736.
Ruaud L, Mignot C, Guet A, Ohl C, Nava C, Heron D et al. DYRK1A mutations in two unrelated patients. Eur J Med Genet 2015; 58: 168–174.
Zhou Z, Hong EJ, Cohen S, Zhao WN, Ho HY, Schmidt L et al. Brain-specific phosphorylation of MeCP2 regulates activity-dependent Bdnf transcription, dendritic growth, and spine maturation. Neuron 2006; 52: 255–269.
Cheng TL, Wang Z, Liao Q, Zhu Y, Zhou WH, Xu W et al. MeCP2 suppresses nuclear microRNA processing and dendritic growth by regulating the DGCR8/Drosha complex. Dev Cell 2014; 28: 547–560.
Hammerle B, Elizalde C, Tejedor FJ . The spatio-temporal and subcellular expression of the candidate Down syndrome gene Mnb/Dyrk1A in the developing mouse brain suggests distinct sequential roles in neuronal development. Eur J Neurosci 2008; 27: 1061–1074.
Penzes P, Cahill ME, Jones KA, VanLeeuwen JE, Woolfrey KM . Dendritic spine pathology in neuropsychiatric disorders. Nat Neurosci 2011; 14: 285–293.
Salichs E, Ledda A, Mularoni L, Alba MM, de la Luna S . Genome-wide analysis of histidine repeats reveals their role in the localization of human proteins to the nuclear speckles compartment. PLoS Genet 2009; 5: e1000397.
Rueda N, Mostany R, Pazos A, Florez J, Martinez-Cue C . Cell proliferation is reduced in the dentate gyrus of aged but not young Ts65Dn mice, a model of Down syndrome. Neurosci Lett 2005; 380: 197–201.
Clark S, Schwalbe J, Stasko MR, Yarowsky PJ, Costa AC . Fluoxetine rescues deficient neurogenesis in hippocampus of the Ts65Dn mouse model for Down syndrome. Exp Neurol 2006; 200: 256–261.
Roper RJ, Baxter LL, Saran NG, Klinedinst DK, Beachy PA, Reeves RH . Defective cerebellar response to mitogenic Hedgehog signaling in Down [corrected] syndrome mice. Proc Natl Acad Sci USA 2006; 103: 1452–1456.
Contestabile A, Fila T, Cappellini A, Bartesaghi R, Ciani E . Widespread impairment of cell proliferation in the neonate Ts65Dn mouse, a model for Down syndrome. Cell Prolif 2009; 42: 171–181.
Contestabile A, Fila T, Ceccarelli C, Bonasoni P, Bonapace L, Santini D et al. Cell cycle alteration and decreased cell proliferation in the hippocampal dentate gyrus and in the neocortical germinal matrix of fetuses with Down syndrome and in Ts65Dn mice. Hippocampus 2007; 17: 665–678.
Guidi S, Bonasoni P, Ceccarelli C, Santini D, Gualtieri F, Ciani E et al. Neurogenesis impairment and increased cell death reduce total neuron number in the hippocampal region of fetuses with Down syndrome. Brain Pathol 2008; 18: 180–197.
Ji J, Lee H, Argiropoulos B, Dorrani N, Mann J, Martinez-Agosto JA et al. DYRK1A haploinsufficiency causes a new recognizable syndrome with microcephaly, intellectual disability, speech impairment, and distinct facies. Eur J Hum Genet 2015; 23: 1473–1481.
Bronicki LM, Redin C, Drunat S, Piton A, Lyons M, Passemard S et al. Ten new cases further delineate the syndromic intellectual disability phenotype caused by mutations in DYRK1A. Eur J Hum Genet 2015; 23: 1482–1487.
Brett M, McPherson J, Zang ZJ, Lai A, Tan ES, Ng I et al. Massively parallel sequencing of patients with intellectual disability, congenital anomalies and/or autism spectrum disorders with a targeted gene panel. PLoS One 2014; 9: e93409.
Yu L, Wu Y, Wu BL . Genetic architecture, epigenetic influence and environment exposure in the pathogenesis of autism. Sci China Life Sci 2015; 58: 958–967.
Guedj F, Pereira PL, Najas S, Barallobre MJ, Chabert C, Souchet B et al. DYRK1A: a master regulatory protein controlling brain growth. Neurobiol Dis 2012; 46: 190–203.
Moller RS, Kubart S, Hoeltzenbein M, Heye B, Vogel I, Hansen CP et al. Truncation of the Down syndrome candidate gene DYRK1A in two unrelated patients with microcephaly. Am J Hum Genet 2008; 82: 1165–1170.
Benavides-Piccione R, Dierssen M, Ballesteros-Yanez I, Martinez de Lagran M, Arbones ML, Fotaki V et al. Alterations in the phenotype of neocortical pyramidal cells in the Dyrk1A+/- mouse. Neurobiol Dis 2005; 20: 115–122.
Fotaki V, Dierssen M, Alcantara S, Martinez S, Marti E, Casas C et al. Dyrk1A haploinsufficiency affects viability and causes developmental delay and abnormal brain morphology in mice. Mol Cell Biol 2002; 22: 6636–6647.
Gwack Y, Sharma S, Nardone J, Tanasa B, Iuga A, Srikanth S et al. A genome-wide Drosophila RNAi screen identifies DYRK-family kinases as regulators of NFAT. Nature 2006; 441: 646–650.
Jiang M, Ash RT, Baker SA, Suter B, Ferguson A, Park J et al. Dendritic arborization and spine dynamics are abnormal in the mouse model of MECP2 duplication syndrome. J Neurosci 2013; 33: 19518–19533.
Acknowledgements
T Dang and WY Duan contributed equally to this study. This work was supported by the 973 program grants from the Ministry of Science and Technology of China: 2013CB945404 and 2010CB529601 (to BLW) and 2011CBA00400 (to ZQ); the Fudan Scholar Award from Fudan University (to BLW); and the CAS Strategic Priority Research Program XDB02050400 (to ZQ). We thank all of the patients and families for their contributions. We are grateful to all members of the 973 project team, especially to Dr Ya-Song Du of Shanghai Mental Health Center and Shanghai Jiaotong University School of Medicine and to Dr Xiu Xu of Children’s Hospital of Fudan University for their contribution to the collection of samples and data for this collaborative project. We thank Dr Yiping Shen, Va Lip, Hong Fang, Chang Lin, Xiaoming Sheng and Hong Shao of the Genetics Diagnostic Lab at Boston Children’s Hospital for their assistance. We thank Gang Li, Renchao Liu, Yanlong Qing and Youhan Yao of the Molecular Diagnostic Center at Children’s Hospital of Fudan University for their assistance.
Author contributions
BL Wu and ZL Qiu conceived, designed and supervised the study. T Dang drafted the manuscript. BL Wu and ZL Qiu revised the manuscript. T Dang performed the neuroscience experiments and analyzed the data. B Yu participated in the IUE assay. DL Tong participated in the slice culture assay. C Cheng participated in the data analysis. YF Zhang prepared the experimental mice. WY Duan, W Wu, K Ye, WX Zhang and M Wu performed the exome sequencing. WY Duan performed the primary analysis of the data and mutation validations. Y An and BB Wu assisted in supervising the study. All authors read and approved the final manuscript.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Additional information
Supplementary Information accompanies the paper on the Molecular Psychiatry website
Supplementary information
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivs 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-nd/4.0/
About this article

Cite this article
Dang, T., Duan, W., Yu, B. et al. Autism-associated Dyrk1a truncation mutants impair neuronal dendritic and spine growth and interfere with postnatal cortical development. Mol Psychiatry 23, 747–758 (2018). https://doi.org/10.1038/mp.2016.253
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/mp.2016.253
This article is cited by
-
Inhibition of Dyrk1A Attenuates LPS-Induced Neuroinflammation via the TLR4/NF-κB P65 Signaling Pathway
Inflammation (2022)
-
Comorbidities associated with genetic abnormalities in children with intellectual disability
Scientific Reports (2021)
-
GABA Signaling Pathway-associated Gene PLCL1 Rare Variants May be Associated with Autism Spectrum Disorders
Neuroscience Bulletin (2021)
-
A Munc18-1 mutant mimicking phosphorylation by Down Syndrome-related kinase Dyrk1a supports normal synaptic transmission and promotes recovery after intense activity
Scientific Reports (2020)
-
Ethanol-Induced Changes in Brain of Transgenic Mice Overexpressing DYRK1A
Molecular Neurobiology (2020)
