
Abstract
Although assist ventilation with FIO2 0.21 is the preferable mode of ventilation in the intensive care unit, sometimes controlled ventilation with hyperoxia is needed. But the impact of this setting has not been extensively studied in elderly subjects. We hypothesized that a high fraction of inspired oxygen (FiO2) and controlled mechanical ventilation (CMV) is associated with greater deleterious effects in old compared to adult subjects. Adult and old rats were submitted to CMV with low tidal volume (6 ml/kg) and FiO2 1 during 3 or 6 h. Arterial blood gas samples were measured at 0, 60 and 180 min (four groups: old and adult rats, 3 or 6 h of CMV), and additionally at 360 min (two groups: old and adult rats, 6 h of CMV). Furthermore, total protein content (TPC) and tumor necrosis factor-alpha (TNF-α) in bronchoalveolar lavage were assessed; lung tissue was used for malondialdehyde and histological analyses, and the diaphragm for measurement of contractile function. Arterial blood gas analysis showed an initial (60 min) greater PaO2 in elderly versus adult animals; after that time, elderly animals had lowers pH and PaO2, and greater PaCO2. After 3 h of CMV, TPC and TNF-α levels were higher in the old compared with the adult group (P < 0.05). After 6 h of MV, malondialdehyde was significantly higher in elderly compared with the adult animals (P < 0.05). Histological analysis showed leukocyte infiltration and edema, greater in old animals. In diaphragm, twitch contraction with caffeine significantly declined after 6 h of CMV only for the elderly group. These data support the hypothesis that relatively short-term CMV with low tidal volume and hyperoxia has greatest impact in elderly rats, decreasing diaphragmatic contractile function and increasing lung inflammation.
Similar content being viewed by others

INTRODUCTION
Aging is associated with anatomical and physiological changes in respiratory system, due to factors as chest wall and spine deformities, emphysema senile, decreased strength of respiratory muscles, decreased vital capacity and increased of residual volume [1]. Moreover, there is deterioration in immunity (termed "immunosenescence") with various insults presumably responsible for the greater susceptibility to infection and decreased capacity of lung repair [2]. Elderly patients have become an increasingly prevalent proportion of the intensive care unit (ICU) population. Historically, the mortality rate for acute respiratory distress syndrome (ARDS) has been 69 % to 80 % among elderly patients, with impact in the health care public services [3]. Although outcomes of patients with ARDS have significantly improved in recent years, the studies on ARDS rarely include substantial numbers of elderly patients.
Although assist ventilation with FIO2 21 % is the preferable mode of ventilation in the ICU [4, 5], sometimes controlled ventilation with hyperoxia is needed [6].
Mechanical ventilation (MV) maintain ventilation when patients cannot sustain a good alveolar ventilation, e.g., during respiratory failure, neuromuscular diseases, drug overdoses, and recovery from general anesthetics [7]. However, MV can be associated with deleterious effects. During controlled MV, the diaphragm is mechanically inactive and rapidly develops atrophy and oxidative injury [8, 9]. Ventilation with high tidal volume causes acute lung injury that leads to changes such as increased microvascular permeability, the influx of neutrophils, the production of plasminogen activator inhibitor-1 and the mRNA expression of plasminogen activator inhibitor-1 [10]. Collectively, the aforementioned studies indicate that prolonged MV impairs diaphragmatic contractile function and increases lung inflammation. However, the impact of MV in elderly human individuals is not well known.
Hyperoxic acute lung injury (HALI) is related to the degree of inspired oxygen fraction (FiO2), its duration and perhaps to genetic background, and generates an excess of oxygen/nitrogen reactive species that could not be handled by the organism and causes cell death — initially there is pulmonary edema, followed by fibroproliferation; HALI mortality is between 25 % and 100 % in primates studies, where respiratory distress took 2–8 days to develop [11]. However, some studies contest the lung toxicity of hyperoxia in humans, after showing no or reversible microscopic differences [12, 13]. Despite the presumable deleterious action of hyperoxia, higher FiO2 can be used to cause redistribution of pulmonary blood flow and to decrease vasoconstriction in atelectatic or non-ventilated areas [14, 15]. The administration of FiO2 of 1.0 is also a standard practice to maximize the oxygen reserve during emergence from anesthesia, and higher FiO2 need to be used in some specific situations as, for example, elective Caesarean section under general anesthesia for transfer oxygen to fetus [16, 17]. These are examples of short use of hyperoxia, but longer uses can be found in the setting of very severe ARDS [6, 11].
The association of hyperoxia and mechanical ventilation with high tidal volume is related to greater alterations than that found when they are used, respectively, with a low tidal volume or a FiO2 of 0.21 [18]. But it has been shown that hyperoxia and mechanical ventilation can induce different lung responses to the resultant increased level of chemokines (protection vs. apoptosis), suggesting that the cumulative effect of various factors can generate complex settings [19]. In this way, aging would be another factor that could interact with hyperoxia and mechanical ventilation in order to creating a setting different from that encountered in adults submitted to the same situation. Along with this hypothesis, it has been shown that aging and mechanical ventilation can generate additive effects [20].
The present study was designed in an animal model to determine whether elderly subjects are more susceptible than adult to the effects — on lung and diaphragm functions and on the accompanying inflammatory response — of two different times of mechanical ventilation (3 and/or 6 h) with low tidal volume and high FiO2.
MATERIALS AND METHODS
Experimental Animals and Study Design
Procedures were approved by the Institutional Research Committee at the Federal University of São Paulo and governmental authorities, and followed the guidelines for animal experiments established by the American Physiology Society.
Healthy, adult (8 months old, n = 10) and elderly (20 months old, n = 10) male Wistar (Rattus norvegicus albinus) rats were individually housed and fed with rat chow (calorie-reduced diet) and water ad libitum and were maintained on a 12:12-h light–dark cycle. The small number of animals was used with base in other studies [21–23], our previous experience [24] and the guidelines of the 3R's (Council Directive 86/609/EEC and New limits for the use of animals in experiments by the European Parliament in 2010). The animals were randomly divided in: adult ventilated for 3 h (AV3; n = 5) and ventilated for 6 h (AV6; n = 5), elderly ventilated for 3 h (EV3; n = 5) and ventilated for 6 h (EV6; n = 5).
Protocol for MV-Treated Animals
All surgical procedures were performed using aseptic technique. After reaching a surgical plane of anesthesia (sodium thiopental, 50 mg/kg [Thionembutal, Abbott, SP, Brazil]; intraperitoneally [ip]), the animals were tracheotomized and a 14G cannula was inserted into the trachea, and the sterile PE50 catheter was inserted into the left internal carotid artery for blood gas sampling. Animals were ventilated via tracheotomy. Muscle relaxation was performed with suxamethonium ip (0.8 mg/kg; União Química Farmacêutica Nacional, Sao Paolo, Brazil), and the animals were immediately connected to a rodent volume controlled ventilator (model 683; Harvard Apparatus, Massachusetts, USA) and ventilated during 3 or 6 h with 100 % FiO2; tidal volume (V T) 6 ml/kg, positive end-expiratory pressure (PEEP) of 5 cmH2O, respiratory rate (RR) of 60–70 cycles/min.
Body temperature was maintained at approximately 37 °C by use of a heating blanket and controlled by rectal thermometer. Animals were constantly monitored by a technician and exposed to continuing care (expressing the bladder, removing upper airway mucus, lubricating the eyes, and passively moving the limbs).
Measurements
Arterial Blood Gas analysis
Blood samples were analyzed for partial pressure of oxygen (PaO2), partial pressure of carbon dioxide (PaCO2) and pH by blood gas analyzer (AVL-Compact3, Roche Diagnostic, Germany) and measured at different times: 0, 60 and 180 min in MV-treated during 3 h. In the groups ventilated for 6 h the blood gas samples were measured at 0, 60, 180 and 360 min.
The animals were euthanized in the end of respective time point (3 and 6 h) using a 1 ml/100 g body weight of the euthanasia solution (T-61; Hoechst & Roussel, SP, Brazil) after diaphragm removal.
Bronchoalveolar Lavage (BAL)
At the end of the experiment, median sternotomy was performed, the trachea was isolated, and the left main bronchus was clamped. A 14G catheter was inserted to the right main bronchus of the animal and 10 ml of cold (4 °C) normal saline was instilled into the bronchoalveolar space of the right lung. The instilled fluid was then harvested by aspiration into the syringe. The procedure was repeated three times in each animal. BAL was then filtered through a sterile gauze and centrifuged at 500×g for 15 min at 4 °C to remove mucus and cells. The supernatant was frozen immediately on dry ice and stored at −80 °C for further analysis.
Total Protein Lung Content
Total protein was measured in 1 ml aliquot of BAL using the method of Lowry and colleagues [25].
Cytokine Assay in BAL
Analyses of tumor necrosis factor (TNF)-alpha levels were quantified by ELISA (enzyme-linked immune sorbent assay) kit specifically for the rat (OptEIA Elisa kits; BD Bioscience Pharmigens, California, USA; limit detection of the test was 5–800 pg/ml). The absorbance of each was read at 450 nm with a microplate reader (Labsystem, Multiskan MCC/340, Finland). Procedures were described in the manufacturer's instruction.
Measurement of Lipid Derived Oxidation Products, Malondialdehyde (MDA)
The upper lobe of the right lung was stored at −80 °C for later analysis of lung tissue MDA level as previous described by Buege and Aust [26] and analyzed in spectrophotometer at 530 ηm (E205D, CELM, SP, Brazil).
Measurement of Contractile Function
Entire diaphragm was excised, before euthanasia, and immediately immersed in carboxygenated (95 % O2–5 % CO2) Kreb's Ringer solution, at ambient temperature, with the following composition (in mM/l): 118.1 NaCl, 3.4 KCl, 0.8 MgSO4, 1.2 KH2PO4, 11.1 glucose, 25 NaHCO3, 2.5 CaCl2, and pH 7.4. Diaphragm muscle strips with a median length of 18 mm (25 % percentile: 16, 75 % percentile: 18) and a median cross-sectional area (CSA) of 0.03 mm2 (25 % percentile: 0.03, 75 % percentile: 0.05) were dissected and attached to a support with two platinum electrode along its length; the upper side of the muscle was connected to an isometric force transducer (Myograph F-60; NARCO Bio-Systems, Texas, USA), and length was adjusted to optimize twitch force. The isometric contractions of the diaphragm bundles were recorded in a chamber that was filled with a temperature-controlled carboxygenated Kreb's Ringer solution (37 °C). The stimulation pulses were applied via the platinum electrodes with an electrical stimulator (S88 Dual Channel Square Pulse Stimulator, Grass–Astro-Med, Massachusetts, USA). The isometric force transducer was coupled to an IBM computer, using bridge amplifier/digital-to-analog converter (PCX 0802/CAD 12/32; Lynx Tecnologia Eletrônica, Sao Paolo, Brazil) and biological data recording system (AqDados 4; Lynx Tecnologia Eletrônica). Experiments were carried out after 20 min of incubation, and initial direct stimulation at 0.2 Hz using supramaximal voltage, 1-ms pulses.
Test of Tetanic Stimulation (Tetanic Peak Force — P o)
We measured isometric forces (N) evoked by direct stimulation at 0.2 (evokes initial twitch contraction [ITC]), 20, 30, 40, 50, 60, 80, and 100 Hz. These different frequencies were produced at 20-s intervals. Values were normalized to the CSA and expressed as N/cm2.
Test with Caffeine
Caffeine (C0750; Sigma, St. Louis, MO, USA) was added to the chamber in a progressive way as bolus injection to the final concentrations: 0.5, 1.0, 1.5, 2.0, 3.0, 4.0, and 32 mmol/l. Each dosage of caffeine was given once the level of maximum contracture was reached, or other way, after 3 min. Twitch peak force (P t) at each caffeine concentration (Pt-c) was measured (N). Values were normalized to the CSA and expressed as N/cm2. Caffeine provokes increase of tension on the isolated rat diaphragm: an increase of force in low frequencies (20 Hz) and a decrease in high ones (50–100 Hz), and partly prevents fatigue [27]. At low doses, caffeine increases twitch from normal depolarization; at higher doses there is contracture, i.e., tension without electrical stimulation [28]. Caffeine acts via the sarcoplasmatic reticulum calcium release channel or ryanodine receptor (RYR) and it is also used in the diagnosis of susceptibility to malignant hyperthermia — caused by mutations in the RYR gene [29].
Morphology
A right middle lung lobe was formalin-inflated and fixed. The tissue was subsequently paraffin-embedded and 4-μm sections were stained with hematoxylin and eosin (H&E). Sections were evaluated by a blinded examiner, using light microscopy coupled to a video camera (Axiolab Standart 2.0 and AxionCam; Zeiss, Jena, Germany).
Statistical Analysis
K-S distance was employed for the normality test. Comparisons between groups were made using unpaired t-test. Multiple comparisons were analyzed by a single-factor analysis of variance (ANOVA), followed by Bonferroni's test and Tukey's test. Statistical analyses were performed using a standard computer software package (GraphPad Prisma, GraphPad Software, San Diego, CA, USA). P values <0.05 were considered significant. Data are presented as mean ± SD.
RESULTS
Arterial Blood Gas Analysis
The PaO2 was initially greater in the elderly rats (groups EV3 and EV6) compared with adult rats (AV3 and AV6) at time 60 min, but decreased in all groups at time 180 min; this decrease was more evident in the elderly when compared with adult rats, both at 180 and 360 min (Table 1).
The PaCO2 increased more in the elderly (EV3, EV6) compared with adult groups (AV3, AV6) at 180 and 360 min (Table 2). The pH was lower in the old (EV3, EV6) compared with adult animals (AV3, AV6) at 180 and 360 min (Table 3).
Protein Leakage in BAL
Pulmonary protein content was higher in the elderly (EV3, EV6) compared with their respective adult groups (AV3, AV6), and in the 3 h groups (AV3, EV3) compared with their counterparts of 6 h of controlled mechanical ventilation (CMV) (AV6, EV6) (Fig. 1a).

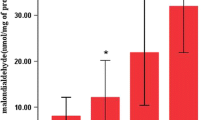
a Effects of mechanical ventilation on total protein content in bronchoalveolar lavage (n = 5 per group). P values of <0.05 were considered statistically significant. AV3 adults ventilated for 3 h, AV6 adults ventilated for 6 h, EV3 elderly ventilated for 3 h, EV6 elderly ventilated for 6 h. b Effects of mechanical ventilation on malondialdehyde (MDA) levels in lung tissue (n = 5 per group). P values of <0.05 were considered statistically significant. AV3 adults ventilated for 3 h, AV6 adults ventilated for 6 h, EV3 elderly ventilated for 3 h, EV6 elderly ventilated for 6 h.
MDA Measurement in the Lung
The MDA levels were higher in the elderly (EV3, EV6) compared with their respective adult groups (AV3, AV6), and in the 3 h groups (AV3, EV3) compared with their counterparts of 6 h of CMV (AV6, EV6) (Fig. 1b).
Cytokine Concentrations in BAL
The TNF-alpha increased more in the elderly (EV3, EV6) compared with their respective adult groups (AV3, AV6), and in the 3 h groups (AV3, EV3) compared with their counterparts of 6 h of CMV (AV6, EV6) (Fig. 2).

Effects of mechanical ventilation on tumor necrosis factor-alpha (TNF-α) levels in bronchoalveolar lavage (n = 5 per group). P values of <0.05 were considered statistically significant. AV3 adults ventilated for 3 h, AV6 adults ventilated for 6 h, EV3 elderly ventilated for 3 h, EV6 elderly ventilated for 6 h.
Measurement of Isometric Contractile Property
There was no significant difference among diaphragm muscle strips length and CSA of the four groups submitted to the tests of tetanic stimulation and caffeine.
Contractile function (N) was evaluated by means of initial twitch (0.2 Hz), tetanic forces after different frequencies stimulation and contraction at each caffeine concentration (Pt-c) for caffeine (Tables 4 and 5). Table 4 shows the ITC at 0.2 Hz and tetanic forces at 20–100 Hz for all four groups. There was no difference in the ITC and in the tetanic forces among all groups (Table 4).
There was no difference in the contraction at each caffeine concentration (Pt-c) between the groups (AV3 vs. AV6, AV3 vs. EV3, and AV6 vs. EV6), but there was a significant decrease of contractility in the EV6 group compared with EV3 group evaluated by means of caffeine concentrations 0.5 to 4 mmol/l (Table 5). After 3 h of mechanical ventilation, the elderly group showed a tendency to a greater diaphragmatic contractility than the adult group, albeit non significant; however, it was lost with the continuation of the experiment.
Morphology
Histological analysis (HE, ×400; Fig. 3) in adult and elderly animals ventilated for 3 h (AV3 and EV3) showed moderate changes in lung, inflammatory cells infiltrate, and neutrophil influx, but these changes were more evident in the elderly animals. The elderly animals ventilated for 6 h (EV6) showed severe changes in lung tissue compared with adult (AV6), and demonstrated massive neutrophil leukocyte infiltration and edema (alveolar, perivascular and interstitial) that resulted in alveolar–capillary wall thickening.

Histopathologic sections of lung. AV3, EV3, AV6, EV6: variable degrees of alveolar, perivascular and interstitial edema, and neutrophils leukocyte infiltration. Hematoxylin and eosin staining. Scale bar: 20 μm.
DISCUSSION
The aging process was studied in our model by means of analysis of three different fields: the lung, the local immune reaction and the muscular activity. Each one has its particular changes in aging, already well studied. But there is not much information about the different ways they react together to mechanical ventilation with hyperoxia.
Regarding lung parenchyma, in particular, with age there is hyperinflation, associated with degeneration of elastic fibers of alveolar duct [30]; however, the lung does not become more compliant, but the chest wall compliance is lower [31]. Other animal studies on the effects of hyperoxia for a long time were associated with lung injury/death after 2–3 days [32]; our results have shown that it is possible that the elderly lung in rat is even more susceptible to hyperoxia. Although humans seem more resistant to the deleterious effects of hyperoxia than rats, high FiO2 induces pulmonary fibrosis with long-term use and atelectasis even with brief use, particularly in the elderly patients [11, 33]. Usually, patients requiring high FiO2, even without lung dysfunction, also receive concurrent MV, with its own positive–negative effects. For avoiding the additional ominous effects of high tidal volume (V T), we have used a low (i.e., 6 ml/kg) rather than a high (12 ml/kg) V T, an approach that has had effects on decreasing mortality in patients with ARDS or ALI (acute lung injury) [4]. In clinical practice, even though the deleterious effects of hypoxia are suspected, moreover with the addictive effect of MV, the effort to avoid hypoxia may predispose clinicians to use a higher FiO2 [34]; it is possible that this approach could be more dangerous to the elderly, which should be studied in the clinical setting of elderly patients with ARDS.
In our study, there was a greater inflammatory response in older rats; it is possible that this reflects both a greater lung lesion and a increase in proinflammatory state in the elderly rat. Models with old rats have also shown a greater ventilation-induced increase in the serum concentrations of aspartate aminotransferase, a nonspecific marker of cell injury, and IL-6, a proinflammatory mediator [35]. The successful aging process is indirectly regulated by cellular and molecular defense mechanisms, such as the degree of DNA repair, apoptosis, immune and neuroendocrine response; these mechanisms lead to a progressive increase in proinflammatory state together with a reduced ability to cope with physiological, chemical and biological stress [36]. In immunosenescence, there is also a decline in the immune response that leads to an increased susceptibility to cancer, infectious diseases and autoimmune diseases [37].
Focusing in muscular activity, aging is independently associated, in skeletal muscle, with loss of mass and strength, greatest susceptibility to injury, atrophy and with less efficient repair; but in normoxia conditions, aging has not exacerbated MV-induced impairment in rat diaphragmatic tension, despite an additive effect [20]. The age of human patients subjected to MV is an independent predictor of ventilator-weaning difficulties [38]. But the effects of MV on aging can be dependent of other factors, such as the length of MV and FiO2. Zergeroglu et al. [39] showed that 3 h of MV did not result in oxidative injury in the diaphragm, which occurred only after 6 h of MV, as indicated by increases in both reactive protein carbonyl derivates and lipid hydroperoxides; moreover, in a model of sepsis, it has been shown that 4 h of MV increases force-generating capacity of the diaphragm, an effect that was evident only in our old group [39, 40]. The decrease of mean maximal force-frequency responses have been described as similar in young and old diaphragms, but with a lower FiO2 (0.21) than that used in our experiment and after a greater time of MV (12 h) [20]. Then, for a shorter time of MV (6 h) and a higher FiO2, our data showed that old diaphragms were more susceptible to impairment than adult ones [7].
The mechanisms accounting for the reduced diaphragm force-generating capacity after controlled mechanical ventilation are multifactorial, but more strongly related to the time of diaphragm inactivity, and include problems in excitation–contraction coupling, myofibril atrophy, decreased myofibril protein concentration, and myofibril injury [41]. Contractility decrease is detected after 12 h in in vitro studies of isometric tension in isolated diaphragmatic strips and seems to be progressive [5]. Eighteen hours of MV is enough to result in reduced diaphragmatic maximal specific tension, but in other way it increases diaphragmatic fatigue resistance [7, 42]. There are even short-term animal models of MV that have shown positive aspects regarding respiratory muscles, such as reversal of respiratory muscle fatigue, prevention of muscle fiber injury, and restoration of perfusion to vital organs in shock [40]. Aging group could be another situation where short term MV with high FiO2 could be beneficial for diaphragm. Moreover, the idea that reactive oxygen species (ROS) are harmful to all cells has been challenged by studies showing that ROS are necessary for contractile function, and levels below or above baseline levels prevent the production of muscle strength [42]. The optimal level of ROS could be reached by use of antioxidants. Li et al. [10], in a study with mice ventilated with high and low tidal volume, showed that treatment with low-dose unfractionated heparin or enoxaparin attenuated the deleterious effects in the lungs of mice ventilated with high tidal volume.
In conclusion, our data indicate that as few as 6 h of MV with high fraction of inspired oxygen can result in decreased diaphragmatic force production, greater lung lesion and increase of inflammatory mediators in aged animals and that the magnitude of these events was time and age-dependent; a greater susceptibility to the adverse effects of mechanical ventilation/hyperoxia in old animals may be relevant to the higher ARDS mortality that has been observed in elderly patients and could prompt researchers to concentrate on specific measures for this particular group. Our observations of the effects of MV/hyperoxia in healthy aged rats should be reproduced in a model that could add a previous lung lesion, for better simulation of the clinical situation of aged patients with ARDS submitted to MV/hyperoxia.
References
Sharma, G., and J. Goodwin. 2006. Effects of aging on respiratory system physiology and immunology. Clinical Interventions in Aging 1: 253––260.
Ostan, R., L. Bucci, M. Capri, S. Salvioli, M. Scurti, E. Pini, D. Monti, and C. Franceschi. 2008. Immunosenescence and immunogenetics of human longevity. Neuroimmunomodulation 15: 224–240.
Eachempati, S.R., L.J. Hydo, J. Shou, and P.S. Barie. 2007. Outcomes of acute respiratory distress syndrome (ARDS) in elderly patients. Journal of Trauma 63: 344–350.
ARDS (The Acute Respiratory Distress Syndrome Network). 2000. Ventilation with lower tidal volumes as compared with traditional tidal volumes for acute lung injury and the acute respiratory distress syndrome. New England Journal of Medicine 342: 1301–1308.
Vassilakopoulos, T., S. Zakynthinos, and C. Roussos. 2006. Bench-to-bedside review: Weaning failure – should we rest the respiratory muscles with controlled mechanical ventilation? Critical Care 10: 204.
Esteban, A., A. Anzueto, F. Frutos, A. Alia, L. Brochard, T.E. Stewart, S. Benito, S.K. Epstein, C. Apeztequia, P. Nightingale, A.C. Arroliga, and M.J. Tobin. 2002. Mechanical Ventilation International Study Group. Characteristics and outcomes in adult patients receiving mechanical ventilation: A 28-day international study. JAMA 286: 345–355.
Powers, S.K., R.A. Shanely, J.S. Coombes, T.J. Koesterer, M. McKenzie, D. Van Gammeren, M. Cicale, and S.L. Dodd. 2002. Mechanical ventilation results in progressive contractile dysfunction in diaphragm. Journal of Applied Physiology 92: 1851–1858.
Betters, J.L., D.S. Criswell, R.A. Shanely, D. Van Gammeren, D. Falk, K.C. Deruisseau, M. Deering, T. Yimlamai, and S.K. Powers. 2004. Trolox attenuates mechanical ventilation-induced diaphragmatic dysfunction and proteolysis. American Journal of Respiratory and Critical Care Medicine 170: 1179–1184.
Shanely, R.A., M.A. Zergeroglu, S.L. Lennon, T. Sugiura, T. Yimlamai, D. Enns, A. Belcastro, and S.K. Powers. 2002. Mechanical ventilation-induced diaphragmatic atrophy is associated with oxidative injury and increased proteolytic activity. American Journal of Respiratory and Critical Care Medicine 166: 1369–1374.
Li, L.F., C.C. Huang, H.C. Lin, Y.H. Tsai, D.A. Quinn, and S.K. Liao. 2009. Unfractionated heparin and enoxaparin reduce high-stretch ventilation augmented lung injury: A prospective, controlled animal experiment. Critical Care 13: R108.
Kallet, R.H., and M.A. Matthay. 2013. Hyperoxic acute lung injury. Respiratory Care 58: 123–140.
Barber, R.E., and W.K. Hamilton. 1970. Oxygen toxicity in man. A prospective study in patients with irreversible brain damage. NEJM 283: 1478–1484.
Davis, W.B., S.I. Rennard, P.B. Bitterman, and R.G. Crystal. 1983. Pulmonary oxygen toxicity. NEJM 309: 878–883.
Lim, D.Y., D.W. Lee, E.A. Jang, S.H. Lee, H.J. Jeong, C.W. Jeong, S.W. Jeong, and K.Y. Yoo. 2012. Effects of inspired oxygen fraction in discriminating venous from arterial blood in percutaneous central venous catheterization under general anesthesia. Korean J Anesthesio. 62: 225–229.
Mackintosh, N., M.C. Gertsch, H.W. Hopf, N.L. Pace, J. White, R. Morris, C. Morrissey, V. Wilding, and S. Herway. 2012. High Intraoperative inspired oxygen does not increase postoperative supplemental oxygen requirements. Anesthesiology 117: 271–279.
Ganter, M.T., C.K. Hofer, and J. Pitter. 2009. Postoperative intravascular fluid therapy. In Anesthesia, 7th ed, ed. R.D. Miller, 2783–2803. Philadelphia: Churchill Livingstone.
Ngan Kee, W.D., K.S. Khaw, K.C. Ma, A.S.Y. Wong, and B.B. Lee. 2002. Randomized double-blind comparison of different inspired oxygen fraction during general anesthesia for Caesarean section. British Journal of Anaesthesia 89: 556–561.
Liu, Y.Y., S.K. Liao, Y.H. Tsai, D.A. Quinn, and L.F. LI. 2009. Role of nuclear factor-κB in augmented lung injury because of interaction between hyperoxia and high stretch ventilation. Translational Research 154: 228–240.
Santos, C.C. 2007. Hyperoxic acute lung injury and ventilator-induced/associated lung injury: new insights into intracellular signaling pathways. Critical Care 11: 126.
Criswell, D.S., R.A. Shanely, J.J. Betters, M.J. McKenzie, J.E. Sellman, D.L. Van Gammeren, and S.K. Powers. 2003. Cumulative effects of aging and mechanical ventilation on in vitro diaphragm function. Chest 124: 2302–2308.
Takeuchi, K., S. Ueki, and S. Okabe. 1986. Importance of gastric motility in the pathogenesis of indomethacin-induced gastric lesions in rats. Digestive Diseases and Sciences 31: 1114–1122.
Gruenewald, D.A., and A.M. Marsumoto. 1991. Age-related decreases in serum gonadotropin levels and gonadotropin-releasing hormone gene expression in the medial preoptic area of the male rat are dependent upon testicular feedback. Endocrinology 129: 2442–2450.
Huang, F.P., G. Xi, R.F. Keep, Y. Hua, A. Nemoianu, and J.T. Hoff. 2002. Brain edema after experimental intracerebral hemorrhage: Role of hemoglobin degradation products. Journal of Neurosurgery 96: 287–293.
Oliveira-Junior, I.S., M.K. Brunialti, I.H. Koh, V.B. Junqueira, and R. Salomão. 2006. Effect of pentoxifylline on lung inflammation and gas exchange in a sepsis-induced acute lung injury model. Brazilian Journal of Medical and Biological Research 39: 1455–63.
Lowry, O.H., N.J. Rosebrough, A.L. Farr, and R.J. Randall. 1951. Protein measurement with the Folin phenol reagent. Journal of Biological Chemistry 193: 265.
Buege, J.A., and S.D. Aust. 1978. Microsomal lipid peroxidation. Methods in Enzymology 52: 302–310.
Gölgeli, A., C. Ozesmi, and M. Ozesmi. 1995. The effects of theophylline and caffeine on the isolated rat diaphragm. Acta Physiologica, Pharmacologica et Therapeutica Latinoamericana 45: 105–113.
Wondmikun, Y., T. Soukup, and T. Asmussen. 2006. Effects of caffeine at different temperatures on contractile properties of slow-twitch and fast-twitch rat muscles. Physiological Research 55: 641–652.
EMHG (European Malignant Hyperpyrexia Group). 1984. A protocol for the investigation of malignant hyperpyrexia (MH) susceptibility. British Journal of Anaesthesia 56: 1267–1269.
Gillooly, M., and D. Lamb. 1993. Airspace size in lungs of lifelong non-smokers: Effect of age and sex. Thorax 48: 39–43.
Mittman, C., N.H. Edelman, A.H. Norris, and N.W. Shock. 1965. Relationship between chest wall and pulmonary compliance with age. Journal of Applied Physiology 20: 1211–1216.
Olivera, W.G., K.M. Ridge, and J.I. Sznajder. 1995. Lung liquid clearance and NaK-ATPase during acute hyperoxia and recovery in rats. American Journal of Respiratory and Critical Care Medicine 152: 1229–1234.
Lumb, A.B. 2007. Just a little oxygen to breathe as you go off to sleep…is it always a good idea? British Journal of Anaesthesia 99: 769–771.
Altemeier, W.A., and S.E. Sinclair. 2007. Hyperoxia in the intensive care unit: Why more is not always better. Current Opinion in Critical Care 13: 73–78.
Nin, N., J.A. Lorente, M. De Paula, P. Fernández-Segoviano, O. Peñuelas, and A. Sánchez-Ferrer. 2008. Aging increases the susceptibility to injurious mechanical ventilation. Intensive Care Medicine 34: 923–931.
Franceschi, C. 1989. Cell proliferation, cell death and aging. Aging 1: 3–15.
Ginaldi, L., M. De Martinis, A. D'Ostilio, L. Marini, M.F. Loreto, M.P. Corsi, and D. Quaglino. 1999. The immune system in the elderly: I. Specific humoral immunity. Immunologic Research 20: 101–108.
Doering, L.V., F. Imperial-Perez, S. Monsein, and F. Esmailian. 1998. Preoperative and postoperative predictors of early and delayed extubation after coronary artery bypass surgery. American Journal of Critical Care 7: 37–44.
Zergeroglu, M.A., M.J. McKenzie, R.A. Shanely, D. Van Gammeren, K.C. DeRuisseau, and S.K. Powers. 2003. Mechanical ventilation-induced oxidative stress in the diaphragm. Journal of Applied Physiology 95: 1116–1124.
Ebihara, S., S.N. Hussain, G. Danialou, W.K. Cho, S.B. Gottfried, and B.J. Petrof. 2002. Mechanical ventilation protects against diaphragm injury in sepsis: Interaction of oxidative and mechanical stresses. American Journal of Respiratory and Critical Care Medicine 165: 221–228.
Sassoon, C.S., V.J. Caiozzo, A. Manka, and G.C. Sieck. 2002. Altered diaphragm contractile properties with controlled mechanical ventilation. Journal of Applied Physiology 92: 2585–2595.
Reid, M.B. 2008. Free radicals and muscle fatigue: Of ROS, canaries, and the IOC. Free Radical Biology and Medicine 44: 169–179.
Acknowledgement
This study was supported by grants from the "Fundação de Amparo a Pesquisa do Estado de São Paulo" (FAPESP: 06/60834-9 and 08/51508-6).
Conflict of interests
The authors declare that they have no competing interests.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Andrade, P.V., dos Santos, J.M., Silva, H.C.A. et al. Influence of Hyperoxia and Mechanical Ventilation in Lung Inflammation and Diaphragm Function in Aged Versus Adult Rats. Inflammation 37, 486–494 (2014). https://doi.org/10.1007/s10753-013-9762-4
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10753-013-9762-4