
Abstract
STUDY QUESTION Is Tcte1 mutation causative for male infertility?
SUMMARY ANSWER Collected data underline the complex and devastating effect of the single-gene mutation on testicular molecular network, leading to male reproductive failure.
WHAT IS KNOWN ALREADY Latest data revealed mutations in genes related to axonemal dynein arms as causative for morphology and motility abnormalities in spermatozoa of infertile males, including dysplasia of fibrous sheath (DFS) and multiple morphological abnormalities in the sperm flagella (MMAF). The nexin-dynein regulatory complex (N-DRC) coordinates the dynein arm activity, and is built from DRC1-DRC7 proteins. DRC5 (TCTE1) – one of N-DRC element, has been already reported as a candidate for abnormal sperm flagella beating, however, only in restricted manner with no clear explanation of respective observations.
STUDY DESIGN, SIZE, DURATION Using CRISPR/Cas9 genome editing technique, mouse knockout line of Tcte1 gene was created on the basis of C57Bl/6J strain. Then, the mouse reproductive potential, semen characteristics, testicular gene expression level, sperm ATP and testis apoptosis level measurements have been performed, followed by visualization of N-DRC proteins in sperm, and protein modeling in silico. Also, a pilot genomic sequencing study of samples from human infertile males (n=248) was applied for screening of TCTE1 variants.
PARTICIPANTS/MATERIALS, SETTING, METHODS To check the reproductive potential of KO mice, adult animals were crossed for delivery of three litters per caged pair, but no longer than for 6 months, in various combinations of zygosity. All experiments were performed for wild type (WT – control group), heterozygous Tcte1+/−, and homozygous Tcte1−/− male mice. Gross anatomy was performed on testis and epididymis, followed by semen analysis. Sequencing of RNA (RNAseq; Illumina) has been done for mice testis tissues. STRING interactions have been checked for protein-protein interactions, based on changed expression level of corresponding genes identified in the mouse testis RNAseq experiments. Immunofluorescence in situ staining was performed to detect the N-DRC complex proteins: Tcte1 (Drc5), Drc7, Fbxl13 (Drc6), and Eps8l1 (Drc3) in mouse spermatozoa. To determine the ATP amount in spermatozoa, the luminescence level was measured. Also, immunofluorescent in situ staining was performed to check the level of apoptosis via caspase 3 visualization on mouse testis samples. DNA from whole blood samples of infertile males (n=137 non-obstructive azoospermia or cryptozoospermia, n=111 samples with spectrum of oligoasthenoteratozoospermia, including n=47 with asthenozoospermia) has been extracted to perform genomic sequencing (WGS, WES or Sanger). Protein prediction modeling of human identified variants and the exon 3 structure deleted in mouse knockout has been also performed.
MAIN RESULTS AND THE ROLE OF CHANCE No progeny at all was found for homozygous males with revealed oligoasthenoteratozoospermia, while heterozygous animals (fertile) manifested oligozoospermia, suggesting haploinsufficiency. RNA-sequencing of the testicular tissue showed the influence of Tcte1 mutations on the expression pattern of 21 genes responsible for mitochondrial ATP processing, linked with apoptosis, or spermatogenesis. In Tcte1−/− males the protein revealed only residual amounts in sperm head nucleus, and was not transported to sperm flagella, as other N-DRC components. Decreased ATP level (2.4-fold lower) was found in spermatozoa of homozygous mice, together with disturbed tail:midpiece ratio, leading to abnormal sperm tail beating. Casp3-positive signals (indicating apoptosis) were observed in spermatogonia only, at similar level in all three mouse genotypes. Mutation screening of human infertile males revealed 1 novel and 5 ultrarare heterogeneous variants (predicted as disease causing) in 6.05% of patients studied. Protein prediction modeling of identified variants revealed changes in the protein surface charge potential, leading to disruption in helix flexibility or its dynamics, thus, suggesting the disrupted TCTE1 interaction with its binding partners located within the axoneme.
INTRODUCTION
Infertility has been defined by World Health Organization as a social disease concerning approximately 10-18% of couples at reproductive age, which are unable to conceive within 1 year of regular unprotected sexual intercourse [WHO, 2020]. Approximately 7% of males and ∼12% of females worldwide reveal reproductive problems, while the male factor is responsible for approximately 20-30% of all infertility cases [Barratt et al., 2017; Vander Borght and Wyns, 2018; Agarwal et al., 2021]. Male infertility is a disease with multifactorial etiology, including: genetic factors, chromosomal aberrations, epigenetic mutations, hormonal abnormalities, infections or reproductive tract abnormalities [Agarwal et al., 2021; Matzuk and Lamb 2008; Zorrilla and Yatsenko 2013; Krausz and Riera-Escamilla 2018]. Genetic causes of male infertility can be detected in ∼10-15% of cases, while in 25-50% of infertile males the etiology remains unexplained [Agarwal et al., 2021; Matzuk and Lamb 2008; Zorrilla and Yatsenko 2013]. The male infertility phenotype mostly results in decreased semen parameters, including: low sperm concentration, abnormal morphology or diminished motility. The actual standards for genetic testing of male infertility rely on chromosomal screening for structural and numerical abnormalities, followed by analyses of deletions in the AZF (azoospermia factor) regions of the Y chromosome [Hwang et al., 2018]. However, it is still not sufficient due to the fact, that approximately 2,000 genes may be related to spermatogenesis, while only ∼10% of them seems to have documented fertility role, so far [Krausz and Riera-Escamilla 2018; Houston et al., 2021] Recent years NGS technology development, including genomic sequencing of DNA (exome or genome; large cohorts of patients), and RNA (RNAseq, scRNAseq on reproductive tissues), followed by animal models of infertility (mostly mouse models; more than 860 genes evaluated, so far [Mouse Genome Informatics, 24 June 2021]) have started the new era of exploration on the massive scale [Houston et al., 2021; Oud et al., 2021; Wyrwoll et al., 2020; Hardy et al., 2021; Xavier et al., 2021].
Beside the genes related to spermatogenesis and low sperm count, also studies of candidates responsible for diminished sperm motility are pending. Thus, the main structural element responsible for motility – the sperm flagella, has to be of high-quality to guarantee sperm motility and then fertilization success. It is known that mutations in genes encoding factors involved in ciliogenesis process, lead to defects in structure and/or functioning of cilia [Girardet et al., 2019; Sironen et al., 2020; Smith et al., 2021; Kumar and Reiter 2021] There is a plenty of data concerning respiratory cilia, however, still little is known about the axonemal dynein arms functioning in sperm tail. Latest data revealed mutations in genes related to axonemal dynein arms as causative for morphology and motility abnormalities in spermatozoa of infertile males [Girardet et al., 2019; Sironen et al., 2020; Aprea et al., 2021; Lorès et al., 2021; Touré et al., 2021; Martinez et al., 2022]. Two definitions were established: (i) for sperm cells with disorganization in axonemal structures, so called dysplasia of fibrous sheath (DFS) [Chemes et al., 1998] and (ii) for multiple morphological abnormalities in the sperm flagella (MMAF), concerning all possible defects of sperm tail, mainly short and thick flagella, and a lack of the central pair (CP) of microtubules or dynein arms [Khelifa et al., 2014]. Spectrum of abnormal sperm tail phenotypes observed allows qualification of a part of disorders into both listed groups (DFS and MMAF) [Sironen et al., 2020; Touré et al., 2021]
The basic structural element of motile cilia and flagella is axoneme built from nine microtubules arranged in doublets positioned radially and one central pair (CP) of singlet microtubules. This central pair is connected to radial spokes (RS) – protein complexes of mechanic regulatory network responsible for regular waveform beating of the cilia [Gui et al., 2019]. RS are linked to inner dynein arms (IDA) localized within each outer dublet. Each outer doublet carries outer dynein arms (ODA) – multi-subunit motor protein complexes, responsible for generation and regulation of sliding between dynein arms leading to ATP-dependent beating [Satir et al., 2014; Wilson et al., 2015]. The 9+2 basic structure is evolutionary conservative [Girardet et al., 2019; Touré et al. 2021; Ishikawa 2017]. Doublets are linked with the nexin-dynein regulatory complex (N-DRC), which coordinates the dynein arm activity and stabilizes the attachment of doublet microtubules. The N-DRC is built from DRC1-DRC7 proteins, among which DRC1, DRC2 and DRC4 are responsible for physical linkage between A and B microtubules of doublets [Gui et al., 2019; Morohoshi et al., 2020; Bowar et al., 2018; Wirschell et al., 2013]. In terms of male infertility, the best studied gene among N-DRC is DRC7 (MIM:618769), which is amenable for proper assembly of the N-DRC structure (in Drc7−/− male mice the length of the sperm tail was reduced, followed by abnormalities of sperm head morphology) [Morohoshi et al., 2020]. Mouse model of Drc6 (Fbxl13) gene (MIM:609080) revealed no essential role in sperm formation and motility [Morohoshi et al., 2020], however, its disruption together with CEP192 (MIM:616426) leads to defects in HEK293T cell motility [Fung et al., 2018]. Mutations in Drc3 gene (MIM:618758) indirectly revealed male infertility as a lack of pregnancy success with wild-type females [Ha et al., 2016].
One of the N-DRC structural genes is TCTE1 (T-Complex-associated-Testis-Expressed 1), known also as DRC5, FAP155, D6S46 in humans (locus 6p21.1; MIM:186975) or Tcte1, D17S, Tcte-1, D17Sil1 in mouse (locus 17 B3; 17 22.54 cM; MGI:98640). TCTE1 is evolutionary conserved in most eukaryotic organisms, that contain motility cilia or flagella in their life cycle (Figure 1), and is required for proper flagella functioning. Expression of TCTE1 is being observed in testis (highest; with expression starting at the stage of spermatid), choroid plexus and Fallopian tube. TCTE1 contains five leucine rich repeats (LRRs). To date, only one report described a role of Tcte1 gene in infertility, where the sperm cells of Tcte1-null mice exhibited asthenozoospermia [Castaneda et al., 2017]. However, understanding of the nature of this specific motility type defect in Tcte1-null animals has been limited.

A Putative homologs of Tcte1 gene conserved in Bilateria, according to HomoloGene (8434), including collation of proteins and their conserved domains; B Sequence alignement between human, mouse and rat; C NCBI Gene schematic representation of Tcte1 gene in Mus musculus.
In our study we aimed to determine the role of Tcte1 in male infertility using mouse knockout model. We created homo- and heterozygous animals, which were next examined for their reproductive potential, differences in anatomy and histology, and semen parameters. We also performed RNA sequencing (RNAseq) in testis tissue to obtain an answer about the influence of Tcte1 null mutation on the expression pattern of other gonadal genes. Also, the ATP measurement of spermatozoa has been performed. Additionally, screening of variants in human samples from males with severely decreased sperm count was made, followed by protein prediction modeling of revealed variants. Thus, this study contains novel and the most complete data concerning TCTE1 role in male infertility, so far.
MATERIALS AND METHODS
Animals
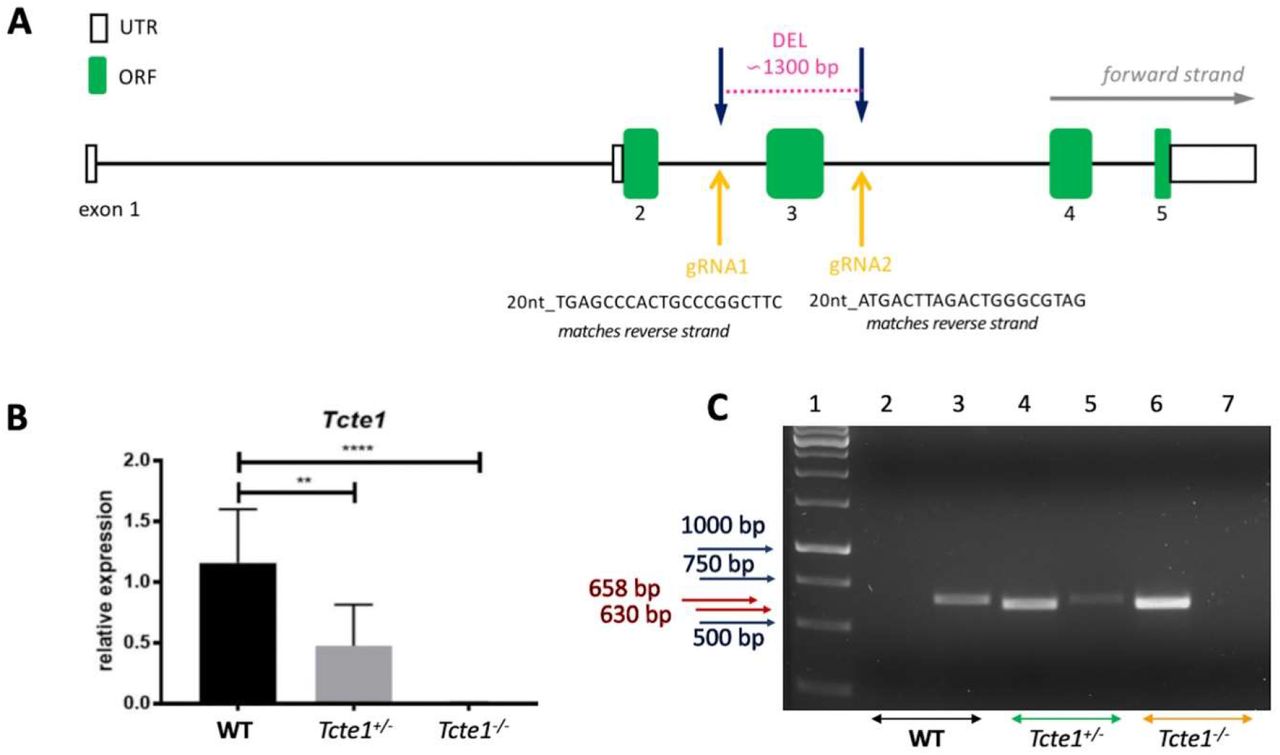
Using CRISPR/Cas9 genome editing technique, mouse heterozygous knockout line of Tcte1 gene (Tcte1+/) was created on the basis of C57Bl/6J strain in the Cyagen Biosciences company (USA). The mouse Tcte1 gene is located on chromosome 17 B3; 17 22.54 cM (GenBank: NM_013688.2; Ensembl: ENSMUSG00000023949). Exon 3 of the Tcte1 gene was selected as target site (deletion of 1292 bp) (Figure 2A). gRNAs targeting vectors (constructed, sequenced and generated by in vitro transcription) were injected into fertilized oocytes (Supplementary file 1). The founders were genotyped and those with positive results were bred to the next generation. Then, in the animal facility of the Institute of Human Genetics PAS (Poznan, Poland), adult Tcte1+/− animals were mated to generate homozygous Tcte1−/− mice. Wild type Tcte1+/+ (WT) animals C57Bl/6J were purchased from Charles River company (Germany). Relative expression of Tcte1 gene was verified using Real-time PCR method (Figure 2B; Supplementary file 2). All mice were housed in pathogen-free animal facility with food and water ad libitum, stable temperature of 22°C, and a light:dark light cycle of 12:12. All procedures were in accordance with the guidelines and regulations for animal experimentations and restricted usage of genetically modified organisms (GMO), and were approved by the Local Ethical Committee for Animal Experiments at the Poznan University of Life Sciences (no. 1/2014, 18/2017), and Ministry of Environment (no. 70/2017). Mouse models with infertility phenotypes were reviewed in the Mouse Genome Informatics database (Jackson Laboratory; http://www.informatics.jax.org/) and through literature searches (PubMed; https://pubmed.ncbi.nlm.nih.gov/).

A Schematic representation of gRNA localization in CRISPR/Cas9 KO creation; B Relative expression of Tcte1 gene in wild type (WT), hetero- (+/−) and homozygous mice (−/−) (Real-time PCR); **** p<0.0001, ** p<0.01; C Example of genotyping result. Lane 1 – 1kb marker (GeneRuler), lanes 2, 3 – wild type (WT), lanes 4, 5 – hetero- (+/−), and lanes 6, 7 – homozygous mice (−/−); lanes: 2, 4, 6 identified the lack (0 bp) or presence of mutation (630 bp), lanes: 3, 5, 7 differentiated hetero- (658 bp) and homozygotes (0 bp).
Reproductive potential
To check the reproductive potential of KO mice, adult animals (7-8 weeks old) were crossed for delivery of three litters per caged pair, but no longer than for 6 months. Crossing combinations included: (i) 4 pairs: homozygous Tcte1−/− male × wild type (WT) female, (ii) 5 pairs: heterozygous Tcte1+/− male × heterozygous Tcte1+/− female, and (iii) 3 pairs: heterozygous Tcte1+/− male × wild type (WT) female. Vaginal plug presence was checked every morning to confirm mating incidence. Time period from mating start point to first pedigree appearance was checked. Number of pups was counted on the day after birth. For each crossed pair, the number of litters, pups, and sex ratio were estimated (also according to pups’ genotypes) (Supplementary file 3).
Genotyping
Genotyping of the pups delivered was performed via PCR assay, with two sets of primers: (i) to identify wildtype and positive KO (het- and homozygous) animals: Tcte1-F: CGTTTTAAGAATGTATTGAGGGTTGGG, Tcte1- R: CTCCGTAGGCTCCTGCCAATATG; estimated product size: for WT 1915 bp, for KO ∼630 bp (deleted ∼1300 bp); PCR results analysis: one band with 1915 bp or 0 bp determined WT, while one band with 630 bp evidenced hetero- or homozygotes; (ii) to distinguish between hetero- and homozygotes: Tcte1-WT/He-F: AGCTTGCCACACCCTCAAGGTACTA, Tcte1-R: CTCCGTAGGCTCCTGCCAATATG; estimated product size: 658 bp for heterozygotes and 0 bp for homozygotes; PCR results analysis: one band with 658 bp determined heterozygotes, while no one band (0 bp) indicated homozygotes (Figure 2C). The PCR mixture and reaction conditions are described in Supplementary file 4.
Tissue gonadal preparation/collection
Gross anatomy was performed on testis and epididymis of WT (n=21), Tcte1+/− (n=22), and Tcte1−/− (n=18) male animals (10 weeks old). Mice were weighted, then euthanized with carbon dioxide (CO2) overdose (euthanasia Minerve Easy-box; Minerve; France) with stable flow rate per minute. Testes and epididymises were collected, weighted, and measured. For histological evaluation tissues were immersed into 10% formalin solution, neutral buffered (Sigma Aldrich, USA) for 3 males per zygosity. Then, Paraplast-embedded sections were prepared (5 µm thickness), using Microtome Leica 2055. To obtain the histological picture, slides were stained with Masson-Goldner protocol [Romeis et al., 2010]. Thereafter, slides were documented with Leica DM5500 light microscope (×1000 magnification, oil immersion).
Mouse semen samples
Mouse spermatozoa were collected from epididymides post mortem. Collected epididymides were placed immediately in a 2.0 mL test tube with warm 500 µL of a non-capacitating Whitten’s HEPES medium (pH 7.2-7.4, 37°C), then cutted into 5-7 fragments, and incubated in 37°C for 15 min. to allow the sperm cells to swim out. Whittens-HEPES medium contained: 100.0 mM NaCl, 4.4 mM KCl, 1.2 mM KH2PO4, 1.2 mM MgSO4, 5.4 mM glucose, 0.8 mM pyruvic acid, 4.8 mM lactic acid (hemi-Ca), HEPES 20 mM [Wertheimer et al., 2013]. Then, spermatozoa were collected and proceeded with seminological analysis (concentration, motility, morphology). Next, semen samples were fixed in a fresh fixative solution (methanol:acetic acid, 3:1 v/v, -20°C; 3×) and stored until further use in -20°C.
Semen analysis
10 μl of sperm suspension was applied on the Makler counting chamber (Sefi medical instrument Ltd., Israel) to test the sperm concentration and total sperm count. Also, the ratio of progressive (fast and slow), non-progressive (total and circular) and immotile spermatozoa were calculated [WHO, 2020]. For assessment of sperm morphology, the Papanicolaou staining was performed. Briefly, the air-dried smears were fixed in ethanol:diethyl ether (1:1 ratio, vol/vol) solution for 10 min. and left for air-drying. Then, slides were stained by Papanicolau procedure according to WHO guidelines [WHO, 2020], and mounted in DPX Mountant (Sigma-Aldrich; USA; cat. no. 06522). A total of 200 spermatozoa were analyzed using Leica DM5500 light microscope (×1000 magnification, oil immersion). Various morphological forms of spermatozoa were determined including abnormalities of head, midpiece and tail. Also, the length of sperm tail, midpiece and the tail:midpiece ratio were measured to check possible differences between males with various zygosity (6 males/per zygosity); for each male at least 100 spermatozoa were measured with the ‘length measurement’ option by CellSense Dimension software (Olympus).
Sequencing of RNA (RNAseq)
For RNA extraction testis tissue was collected into in RNAlater buffer (Invitrogen; Waltham, MA, USA) and freezed immediately (−80°C). Samples were processed using the AllPrep DNA/RNA/Protein Mini Kit (Qiagen; Hilden, Germany). Quality and quantity of extracted RNA samples were checked using NanoDrop (Thermo Scientific) and Quantus (Promega), and 1 μg of RNA per sample has been used for sequencing (Macrogen; South Korea). RNA sequencing was performed with Illumina NovaSeq 6000 platform using TruSeq Stranded mRNA LT Sample Prep Kit, with specified conditions of paired-end reads at 100 bp, and throughput 100 M reads per sample. RNA-Seq raw reads were aligned to mouse reference genome mm10 using hisat 2.0.5 with Gencode vM13 transcriptome reference. Reads aligned in the coding regions of the genome were counted using Feature Counts. Then, read counts were normalized using DESeq2 and subjected to differential analysis. The differential analysis included computing of median based fold change, statistical significance using the Students’ t-test and false discovery rates (correction for multiple testing) in the R/Bioconductor programming environment. Testis tissue expression was retrieved from GTEx (https://gtexportal.org/home/), BioGPS (http://biogps.org/), and AceView (https://www.ncbi.nlm.nih.gov/IEB/Research/Acembly/). All RNA-seq expression data have been deposited in Gene Expression Omnibus (GEO), accession number GSE207805.
STRING interactions
For analysis of protein-protein interactions, based on changed expression level of corresponding genes identified in the mouse testis RNAseq experiments, the STRING analysis was performed [version 11.0]. STRING database allows to delineate probability of the interactions between evaluated proteins using different evidence channels [Mering et al., 2005; Szklarczyk et al., 2019]. This tool enables the integration of all data available in public resources concerning protein-protein interactions.
Immunofluorescence on sperm – N-DRC visualization
Immunofluorescence in situ staining was performed to detect the N-DRC complex proteins: Tcte1 (Drc5), Drc7, Fbxl13 (Drc6), and Eps8l1 (Drc3) in mouse spermatozoa. Mouse spermatozoa were collected and fixed as described above. Specific antibodies were applied: primary antibodies – rabbit anti-human-TCTE1, rabbit anti-mouse-Drc7, anti-mouse-Fbxl13, and anti-mouse-Eps8l1 (all from Biorbyt; Cambridge, UK); secondary antibody – goat anti-rabbit-AF594 (Abcam; cat. no. ab150160; Cambridge, UK). The UniProt IDs and homology of the immunization regions of primary antibodies between human and mouse were described in Supplementary file 5. Antibodies were diluted in 0.5% PBST (primary: 1:100, secondary: 1:500). First, slides with fixed sperm smears were washed in a series of washes in 1× PBST, and then incubated in 25 mM DTT/1 M Tris-HCl, pH 9.5, at room temperature for 20 min. Then, washing in 1× PBST, and incubation in 6 N HCl for 30 min. was applied, followed by blocking step with 1% BSA/1× PBST for 30 min. Next, overnight incubation with antibodies was performed at 4°C in a humidified container. After washing, the samples with 1× PBST, secondary AF594-conjugated antibody was applied for 1 hr. Next, unconjugated antibodies were washed out (4× in 1× PBST, 5 min each, slight rotation). For the final detection, 20 ul of Fluoroshield with DAPI (Sigma-Aldrich; cat. no. F6057) was added to the samples, and microscopic analysis was performed. Images were acquired using a fluorescence microscope with a proper filter set: Leica DM5500, filters: DAPI/TxR/Triple; objectives: 10× and 63× with oil immersion; DFC 7000T camera; software: LASX.
Sperm ATP measurement
For measurement of ATP in spermatozoa the CellTiter-Glo 2.0 Assay (Promega; USA) was used. This ready-to-use assay allows to determine the quantitation of the ATP amount, indicating the presence of metabolically active cells. The assay is based on the thermostability of luciferase (Ultra-GloTM Recombinant Luciferase), and was performed as described in manufacturers’ protocol. Briefly, all laboratory work was done in the stable temperature of 22°C. After collection of mouse spermatozoa (as described above), the amount of approximately 1× 106 of sperm cells was transferred to black opaque 1.5 mL test tube and proper amount of CellTiter-Glo kit was added (1:1, v:v). Next, 2 min. of orbital mixing of the content was performed to activate cell lysis. Then, after 10 min. of incubation (to stabilize the luminescence), the measurement of luminescent signal was completed using Glomax 20/20 luminometer (Promega; USA). For each sample, three measurements (repeats) were done, each in two time points of 0 and +60 min. Numbers of animals used were: WT (n=7), Tcte1+/− (n=7), and Tcte1−/− (n=6). Mean luminescence values obtained (represented as RLU – relative luminescence unit) were compared between tested mice groups.
Immunofluorescence on testis – apoptosis evaluation
Immunofluorescence in situ staining was performed to check the level of apoptosis via caspase 3 visualization on mouse testis samples from all the three zygosities. Caspases play important role in the induction, transduction, and amplification of intracellular apoptotic signals, and thus, caspase-3 can be used as one of the apoptotic indicators [Lej et al, 2022]. Mouse testes were collected and fixed as described above. Specific antibodies were applied: primary antibody – rabbit anti-human-Casp3, (Biorbyt; Cambridge, UK), diluted 1:100 in 0.5% PBST; secondary antibody – goat anti-rabbit-AF594 (Abcam; Cambridge, UK), diluted 1:500 in 0.5% PBST. First, slides with paraffine fixed testis cross sections were deparaffinized in 3 changes of xylene (10 mins. each), and incubated in a series of ethanol: 100%, 96%, and 70%, twice for 5 mins. in each solution. Next, after rinsing in distilled water, tissue sections were incubated sequentially in: a freshly prepared 2% NaBH4/1× PBS, and in 0.1M glycine (30 min., room temp. each). Heat-induced epitope retrieval was performed by immersing the slides in a 0.01M sodium citrate/0.05% Tween 20, pH 6.0 for 30 min., followed by cooling down in a room temp. for next 20 min. Blocking step was done in an aliquot of freshly prepared 1% BSA/1× PBST, at 37 °C for 1 h. Then, tissue sections were incubated with primary antibody in a humidified container overnight at 4°C. Secondary antibody was applied for 1 hr, followed by the washing out of the unconjugated antibody (4× in 1× PBST, 5 min each, slight rotation). For the final detection, 20 μl of Fluoroshield with DAPI (Sigma-Aldrich; cat. no. F6057) was added. Images were captured using a fluorescence Leica DM5500 microscope with a proper filter set: DAPI/SpO/Triple; objectives: 10× and 63× with oil immersion; DFC 7000T camera, LASX software. The analysis was performed for three independent males per zygosity, 2 cross-sections each time, 15-20 tubules per tissue section.
Sequencing of the human samples
A pilot genomic sequencing study of samples from human infertile males was also applied. DNA from whole blood samples (collected into sterile single test tubes with EDTA) was extracted using standard protocols of MagCore HF16 Automated Nucleic Acid Extractor (RBC Biosciences, Taiwan), or Gentra Puregene kit (Qiagen, USA). Total number of sequenced patients was n=248, including: 137 with non-obstructive azoospermia (lack of spermatozoa in ejaculate) or cryptozoospermia (spermatozoa detectable in ejaculate after centrifugation), followed by 111 samples with spectrum of seminal abnormalities in concentration, and/or motility, and/or morphology. Among those samples, 47 revealed decreased motility (asthenozoospermia). Semen evaluation was done according to WHO guidelines [WHO, 2020], followed by AUA and ASRM guidelines for infertile male diagnosis [Schleger et al., 2021]. All men revealed normal hormonal levels, had no chromosomal abnormalities and a lack of AZF microdeletions. For whole exome samples (n=76 from Pittsburgh), sequencing libraries were constructed using the Agilent technologies (Santa Clara, USA), including: SureSelect XT All Exon V4 + UTRs capture kit and SureSelect XT Target Enrichment System. Libraries were sequenced with average read coverage of 62-100x (∼6Gb per sample), using 100 bp paired-end sequencing with TruSeq PE Cluster Kit v3 cBot HS and 4 × 50 TruSeq SBS V3 HS sequencing kit (Illumina, San Diego, USA) on the HiSeq2000 (Illumina, San Diego, CA). Whole genome sequencing (n=42, Poznan) was performed using Illumina HiSeqX with the coverage of at least 30x (100-120 Gb per sample). Sanger sequencing for all exomes of TCTE1 gene (n=130, Poznan) was performed using LightRun96 system of Eurofins Genomics (Eurofins Genomics AT GmbH, Vienna, Austria). Samples were prepared with QIAquick Gel Extraction (cat. no. 28704) or PCR Purification (cat. no. 28104) kits (Qiagen, USA), and analyzed with CLC Main Workbench 8 software (Qiagen, USA).
Filtering of whole genome sequencing results
The quality of raw NGS data was evaluated using the FastQC and MultiQC packages. Reads were aligned to the reference human genome GRCh37 using bwa-mem aligner included in the Speedseq package [Chiang et al., 2015]. For calling of single nucleotide variants (SNVs) and small indels the freeBayes v0.9.21 (speedseq-var) was implemented, following previously described pipeline [Supernat et al., 2018]. Variant disease association and missense mutation prediction effects were determined using: dbSNP (https://www.ncbi.nlm.nih.gov/snp/), OMIM (https://www.ncbi.nlm.nih.gov/omim/), ClinVar (https://www.ncbi.nlm.nih.gov/clinvar/), MutationTaster (http://www.mutationtaster.org/), SIFT (https://sift.bii.a-star.edu.sg/), Polyphen (http://genetics.bwh.harvard.edu/pph2/), CADD (https://cadd.gs.washington.edu/snv), MetaDome (https://stuart.radboudumc.nl/metadome/), REVEL (rare exome variant ensemble learner) [Ioannidis et al., 2016], Ensembl Variant Effect Predictor [Mclaren et al., 2016], and Human Gene Mutation Database (HGMD; Qiagen, Redwood City, CA). SNV pathogenicity was assessed on the basis of American College of Medical Genetics and Genomics (ACMG) guidelines [Richards et al., 2015]. PhyloP (Cold Spring Harbor Laboratory) and Clustal Omega were used for determination of conservation (https://www.ebi.ac.uk/Tools/msa/clustalo/). All NGS variants were confirmed via the Integrative Genomics Viewer (Broad Institute). Variant frequencies have been obtained from the gnomAD database (frequencies from more than 125k exomes and 15k genomes, http://gnomad.broadinstitute.org; v2.1.1), and were filtered in search of rare variants (with minor allele frequency (MAF) <1%).
Protein prediction
The amino acid sequence of TCTE1 was input into Phyre2 [Kelley et al., 2015] for both intensive and fast mode homology modeling, I-TASSER [Roy et al., 2010; Yang et al., 2015, Yang and Zhang 2015], and AlphaFold [Jumper et al., 2021; Varadi et al., 2022] which utilize sequence alignment-based algorithms for generating predicted secondary and tertiary structures; AlphaFold additionally uses machine learning to integrate biological principles of protein folding with the alignment-based algorithms. Confidence scores were obtained from these programs; AlphaFold additionally generates predicted aligned error plots for assessing the quality of the model’s long-range interactions. Models were inspected manually and aligned using the secondary structure matching (SSM) [Krissinel and Henrick, 2004] tool in COOT [Emsley et al., 2010] to minimize user input bias. Docking of the structures into electron tomography data was performed manually in COOT [Emsley et al., 2010]. No refinement was performed to minimize bias. All structure model figures and mutations without post refinement, were generated using Pymol (Schrödinger). I-TASSER was used also for prediction of the exon 3 structure deleted in our mouse KO model (Supplementary file 6).
Statistical analysis
Statistical analyses included the following: normality test (D’Agostino and Pearson or Kolmogorov-Smirnoff), unpaired two-tailed t-test with Welch’s correction, one-way ANOVA test and Fisher’s exact, and two-tailed Pearson correlation. All tests were performed with a significance level of α=0.05 using GraphPad Prism (v.7.0e) software.
RESULTS
Reproductive potential of KO mice
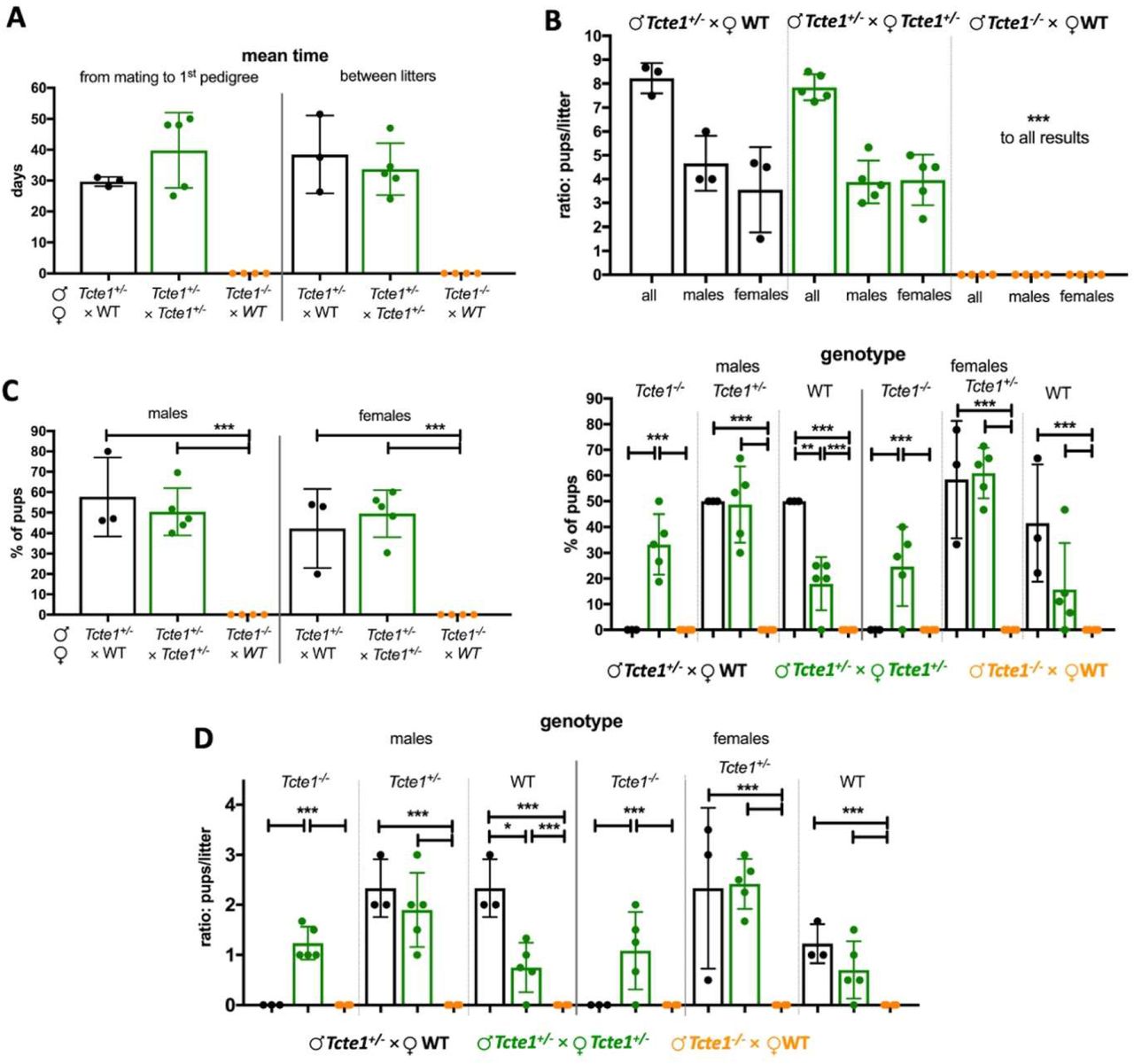
Reproductive potential results were shown in Figure 3 and Supplementary file 3. Only pairs with homozygous Tcte1−/− males revealed no progeny (Tcte1−/− male x WT female). Other combinations (Tcte1+/− male × Tcte1+/− female; Tcte1+/− male × WT female) showed similar results, when considering: number of litters, pups, and sex ratio. Mean time of caging to delivery of the 1st pedigree was longer in Tcte1+/− male × Tcte1+/− female combination by 25.45% (statistically not significant). When considering genotypes of the pups, statistically significant differences (p<0.05) were observed in the number of Tcte1−/− male and female pups delivered – no homozygous animals were obtained from combination of Tcte1+/− male × WT female, while in mating of Tcte1+/− male × Tcte1+/− female, homozygous males (33.25% of all pups) and females (24.67%) were received.

KO Tcte1 mice. Mating combinations and their results including: time of breeding, number of litters and pups, number of male and female pups, and genotypes observed in pups of each combination. A Mean time: from mating to 1st pedigree, and between particular litters; B Ratio of males and females per litter in each mating combination; C Frequency of males and females and their genotype observed each mating combination; left panel: according to sex of pups; right panel: according to mating combinations; D Ratio of males and females per litter and their genotype observed each mating combination. *** p < 0.0001, ** 0.0001 < p < 0.01, * 0.01 < p < 0.05
Histopathology and seminal parameters
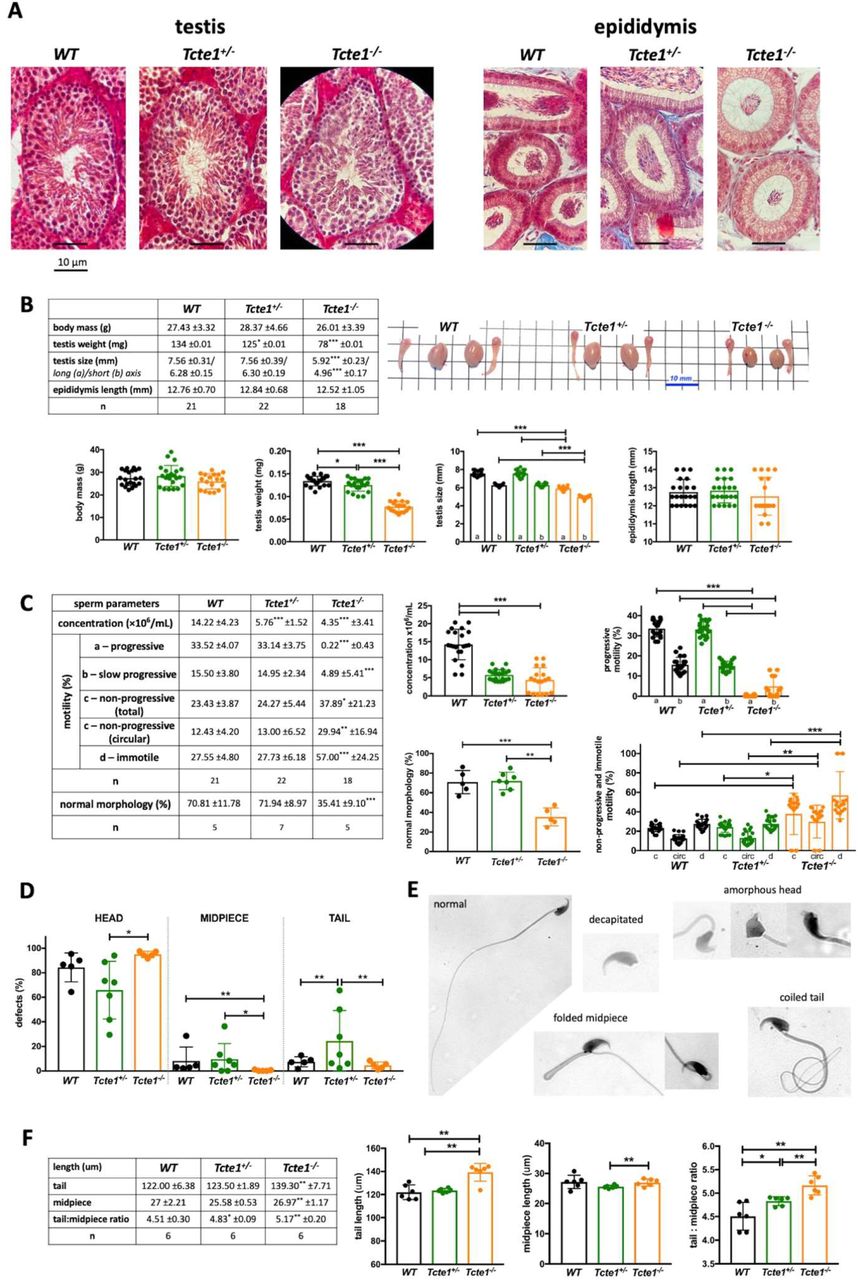
Histopathological evaluation of testicular and epididymal sections revealed no differences between WT, Tcte1+/−, and Tcte1−/− males (Figure 4A). No odds were found for body mass of the animals and the length of the epididymis (Figure 4B). In the Tcte1−/− males testis weight was decreased by approximately 41% (p<0.0001) followed by the ∼22% decline of its size (p<0.0001), while in Tcte1+/− males only testis weight was slightly decreased (approx. 7%; p<0.05) when compared to control animals.

A Histopathology of testes and epididymises (caput); staining: Masson-Goldner protocol; microscope: Leica DM5500, objective 63×, magnification 630×; B Comparison of body mass, testis weight and size, and epididymis length, followed by gross morphology of gonads; C Sperm parameters: concentration, motility and morphology comparison; statistical marks indicated differences according to WT animals; D Frequency of morphological defects according to three part of the spermatozoa; E Examples of main sperm morphological abnormalities found (all abnormalities observed available in Supplementary file 7); F Comparison of the mean length of: sperm tail, midpiece and ratio tail:midpiece; *** p < 0.0001, ** 0.0001 < p < 0.01, * 0.01 < p < 0.05
When considering seminal parameters, both: in Tcte1−/− as well as in Tcte1+/− males the sperm concentration was importantly decreased (3.27, and 2.47-fold, respectively), when collating to control results (p<0.0001; Figure 4C). Additionally, in Tcte1−/− males the frequency of progressively motile spermatozoa was residual, with progressive type of motility in only 0.22% of spermatozoa (decreased ∼150-fold vs. control 33.52%, (p<0.0001)), followed by 4.89% of slow progressive spermatozoa (decreased ∼3-fold vs. control 15.50%) (p<0.0001). Interestingly, ∼2.4-fold increase in circular type of motility (around own axis and non-progressive) was observed for Tcte1−/− males when compared to WT and Tcte1+/− animals (p<0.01) (Supplementary file 7; Supplementary file 8; Supplementary file 9). The frequency of immotile spermatozoa also increased in Tcte1−/− males (2-fold to control; p<0.0001). The frequency of morphologically normal spermatozoa in Tcte1−/− animals was significantly decreased (2-fold to control; p<0.0001; Figure 4C, D, E; Supplementary file 10). The majority of morphological defects was observed for sperm head, mostly of decapitated spermatozoa in Tcte1−/− animals – 52.55% vs. 19.16% for Tcte1+/− and 23.33% for WT, and amorphous head shape with decreased frequency in Tcte1−/− (26.11%) and Tcte1+/− (33.72%) males vs. WT animals (48.88%) (Supplementary file 10). Midpiece defects were found in Tcte1+/− (9.71%) and WT (8.20%) animals, while Tcte1−/− mice revealed only low frequency (0.38%) of such defects. On the other hand, an increased frequency (∼5-fold) of coiled tail was observed for Tcte1+/− animals when compared to other mice zygosity (p<0.01) (Supplementary file 10). Summarizing, seminological changes in mice with Tcte1 mutation resulted in oligoasthenoteratozoospermia in homozygous Tcte1−/− or oligozoospermia in heterozygous Tcte1+/− animals.
Increased mean length of tail and tail:midpiece ratio in Tcte1−/− animals (139.30 ± 7.71 um; 5.16, respectively) was measured when comparing to WT (122.00 ± 6.375 um; 4.51; p<0.002) and Tcte1+/− (123.50 ± 1.89; 4.83; p<0.008) (Figure 4F). Also, in the Tcte1+/− the midpiece length differed from Tcte1−/− animals (p=0.0334), followed by tail:midpiece ratio differences when compared to WT (p=0.0435).
Sequencing of RNA (RNAseq) and STRING pathway analysis
RNA sequencing analysis of gene expression in testes of KO mice revealed statistically significant changes (p<0.05) in 21 genes, including 5 genes both for Tcte1−/− vs. Tcte1+/− and WT, 8 genes only for Tcte1−/− vs. WT, 7 genes only for Tcte1−/− vs. Tcte1+/−, and 1 gene both for Tcte1+/− vs. Tcte1−/− and WT (Figure 5). In Tcte1−/− animals vs. both: Tcte1+/− and WT, the expression level of 4 genes from kallikrein subfamily was decreased 2-8-fold, including genes involved in semen liquefaction and degradation of extracellular matrix proteins in the interstitial area surrounding Leydig cells of the adult mouse testes (Klk1b27, Klk1b21, Klk1b24), or neuronal activity (Klk1b22) (Figure 5; Supplementary file 11) [Michaelis et al., 2017; Marques et al., 2016; Du Plessis et al., 2013; Fink et al., 2007]. 2-fold decrease of expression of genes related to mitochondrial cytochrome oxidase subunits and asthenozoospermia was documented (mt-Co2, mt-Co3) (Figure 5) [Heidari et al., 2016; Mostafa et al., 2016; Baklouti-Gargouri et al., 2013]. In a scheme: Tcte1−/− vs. WT, 8 genes revealed changes in expression level, including 4.7-fold rise for Fetub gene, which is known as required for egg fertilization (via specific inhibitor activity of the zona pellucida (ZP) ovastacin, the cysteine protease responsible for ZP2 cleveage) [Binsila et al., 2021; Karmilin et al., 2019], and plays also a role of a secreted factor during reprogramming [Bansho et al., 2017].

Volcano plots: black dots indicated all analyzed genes within the genome, red dots represented genes with significantly changed expression level (followed by the heatmap and the table). Filtering criteria: p val <0.05, log ratio >1 or <-1 (adequate to fold change value of 2 (expression level increased) or 0.5 (expression level decreased)), expression level >8. nd – no data concerning function or disease.
Two-fold rise for Igf2 gene (strongly linked to spermatogenesis, sex determination, testis development, infertility and prostate cancer; [Matsuzaki et al., 2020; Cannarella et al., 2020; Neirijnck et al., 2019; Damaschke et al., 2017]) has been also noticed. Igf2 and Klk proteins are enclosed in a common network consisted of proteins directly linked to fertility (Figure 6; Supplementary file 11; Supplementary file 12). One of them is Ambp, a fusion protein, which proteolysis generates bikunin – a protein which deficiency results in infertility, due to the diminished formation of the stable cumulus-oocyte complex required for maturation and ovulation [Sato and Kan,2001; Naz and Rajesh, 2005]. Bikunin is one of the elements of ITIH domains (Inter-α-Trypsin Inhibitor Heavy-chain), responsible for hyaluronian metabolism [Hull et al., 2012; Salustri et al., 2019]. ITIH is also linked to Plg, which has the protease activity and activates metalloproteins, such as: Mmp8 and Mmp9, also included in the network. Mmp8 is engaged in degradation/stabilization of occluding and blood-testis barrier [Chen et al., 2018], while Mmp9 (with Igf1 and Igf2), is important for normal placentation [Ghosh et al., 2011]. Igf1 and Igf2, are related to Igfbp3, which interacts with humanin expressed in Leydig cells. Humanin together with IGFBP3 and Bax prevents the activation of germ cell apoptosis [Jia et al., 2015; Lue et al., 2021]. What is important, expression of IGFBP3 is regulated by HOXD10 gene [Xue et al., 2013; Pan et al., 2021], of which expression has been found with 2.9-fold rise in our study. Hoxd10 is involved in prostate cancer processing, cell differentiantion and morphogenesis [Jonkers et al., 2020; Mo et al., 2017; Zhang et al., 2019].

STRING analyzis of potential interactions between proteins with observed differences in gene expression level in RNAseq for mouse testis of the KO model for Tcte1 gene. Genes revealed in RNAseq were bracketed. (STRING database, 24th Feb 2021).
Another gene with 2.2-fold expression rise is Slc45a3, a well-known marker of prostate cancer progression, when a loss of the protein determined poorer prognosis for cancer patients [Hernández-Llodrà et al. 2017; Pin et al., 2017; Perner et al., 2013]. An overexpression of another gene identified in our study (12-fold rise for Usp39 gene) is linked with promotion of prostate or ovarian tumorigenesis. Usp39 is also involved in pre-mRNA splicing, cytokinesis and spindle checkpoint working, and is crucial for apoptosis [Huang et al., 2016; Yuan et al., 2021; Zhao et al., 2016].
Also, 2.2-2.5-fold rise for ncRNA genes (Gm26848, Gm45648; no data concerning their role) and 2-fold reduction for one pseudogene (Gm28661; also no data) were found (Figure 5). Following the latest data (November 2021) from Gene NCBI database, the Gm45648 has been annotated as one with the testis-restricted expression in mouse but no function has been suggested. The second ncRNA Gm26848 found is suggested as protein coding organic cation transporter-like protein across membranes in Drosophila sechellia. There were also two other genes with expression change (3.6-fold decrease – pseudogene Gm44126, and 2.5-fold increase – 4933430L12Rik probably involved in antisense transcription). ‘Gene’ database data showed that the Gm44126 in mice is rather a protein coding gene with the function of UDP-glucuronic acid decarboxylase, and contains two domains involved in NAD(P)H processing. Thus, it can be suggested that this gene is as a novel candidate involved in mitochondrial respiratory chain. Our data with 3.6-fold expression decrease of this gene seem to fit into observed diminished functioning of mitochondrial machinery in Tcte1−/− mice.
Another three genes manifested 2-fold decrease of their testis expression when comparing Tcte1−/− vs. Tcte1+/− (but not to WT): Aqp2 (aquaporin-2 involved in osmotic transportation of the water), with well-known role in spermatogenesis, female endometrium cycle and urinary incontinenece; [Klein et al., 2013; Ramli et al., 2019], Mt3 (metallothionein-3, committed in homeostasis and detoxification of toxic metals, also in testis) important in apoptosis regulation of Bax and Caspase pathways [Gao et al., 2021; Honda et al., 2010]), and Rps27rt (ribosomal protein) (Figure 5; Supplementary file 11). It was also found that Rps27rt may interact indirectly with Usp39 and mitochondrial respiratory complex linking the mitochondrial energy processing with apoptosis (Supplementary file 11). Last gene – Ube2l6, was found to have 2-fold increased expression level in testis in Tcte1+/− animals vs. Tcte1−/− and WT. Its role has been established as ubiquitin enzyme engaged in: stress damage response, cell cycle progression, embryo development, immune response, and factors linked to ATP machinery (Figure 5) [Huang et al., 2016; Yuan et al., 2021; Zhao et al., 2016; Sandy et al., 2020; Cai et al., 2021]. Increased expression of Ube2l6 was linked to degradation/suppression of cancer cells and affected downstream apoptotic factors (i.e., Caspase 3, Caspase 9, Bcl-2, Bax) [Li et al., 2018].
Immunofluorescence on sperm
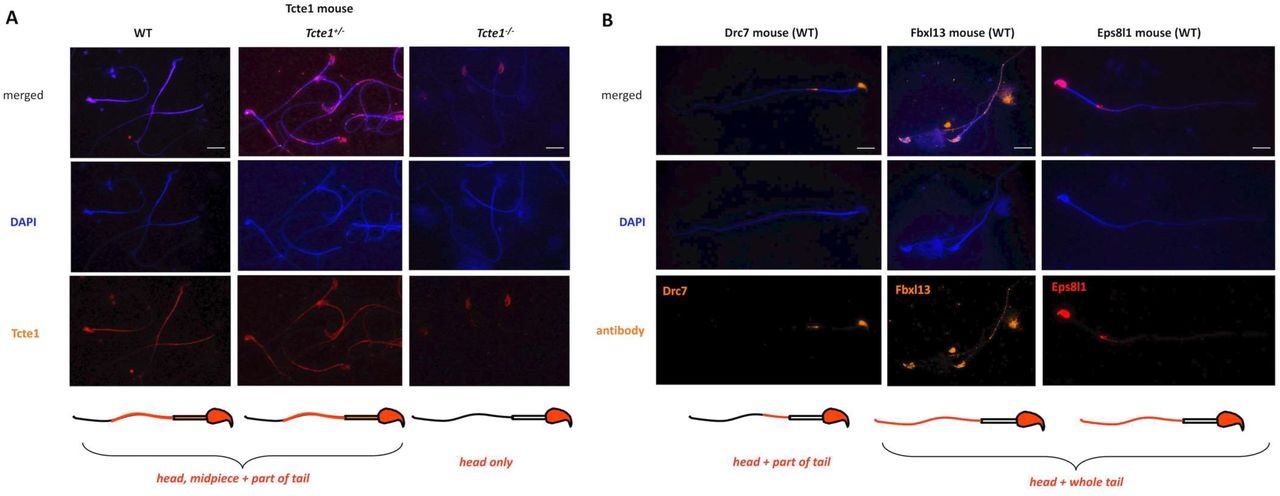
An example of immunofluorescence in situ staining on mouse spermatozoa used to detect the N-DRC proteins has been presented in Figure 7. WT mouse spermatozoa showed Tcte1 protein presence in the sperm head nucleus, midpiece, and a part of the tail (Figure 7A). Immunofluorescent signal of Tcte1 in Tcte1+/− mice was similar to WT animals, while in Tcte1−/− was localized only in the nucleus (Figure 7A). It seems to confirm that in Tcte1−/− males the protein has not been produced properly, revealing only residual amounts in sperm head nucleus, and not transported to sperm flagella. The other N-DRC components (Drc7, Fbxl13 and Eps8l1) were localized in the sperm nucleus and among the whole sperm tail (Fbxl13, Eps8l1) or only within its fragment (Drc7), with exception of the midpiece (Figure 7B). Results obtained seem to suggest co-localization of evaluated N-DRC proteins in various combinations, not common among the midpiece and the terminal part of the sperm tail.
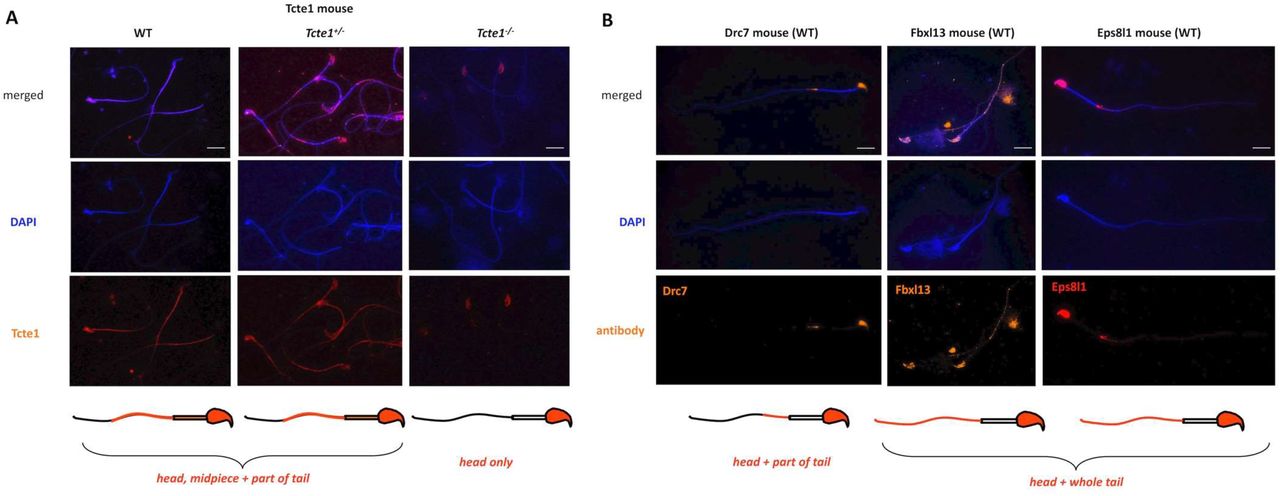
Localization of Tcte1 in spermatozoa from wild type (WT), hetero- (+/−) and homozygous (−/−) animals; B. Localization of other proteins (Drc7, Fbxl13, Eps8l1) building the nexin-dynein regulatory complex (N-DRC), responsible for coordination of the dynein arm activity and stabilization of the doublet microtubules attachment; wild type (WT) spermatozoa. Antibodies: primary (all anti-rabbit 1:100; Biorbyt, UK): Tcte1 (orb357083); Drc7 (orb58695); Fbxl13 (orb678278); Eps8l1 (orb382538); secondary: goat anti-rabbit-AF594 conjugated, 1:500, ab150160, Abcam, UK. Fluorescent microscope: Leica DM5500, filters – DAPI, TxR, FITC, BGR, magnification 630× (with immersion); software: LASX or CytoVision. Bar represents 10 μm.
Sperm ATP level
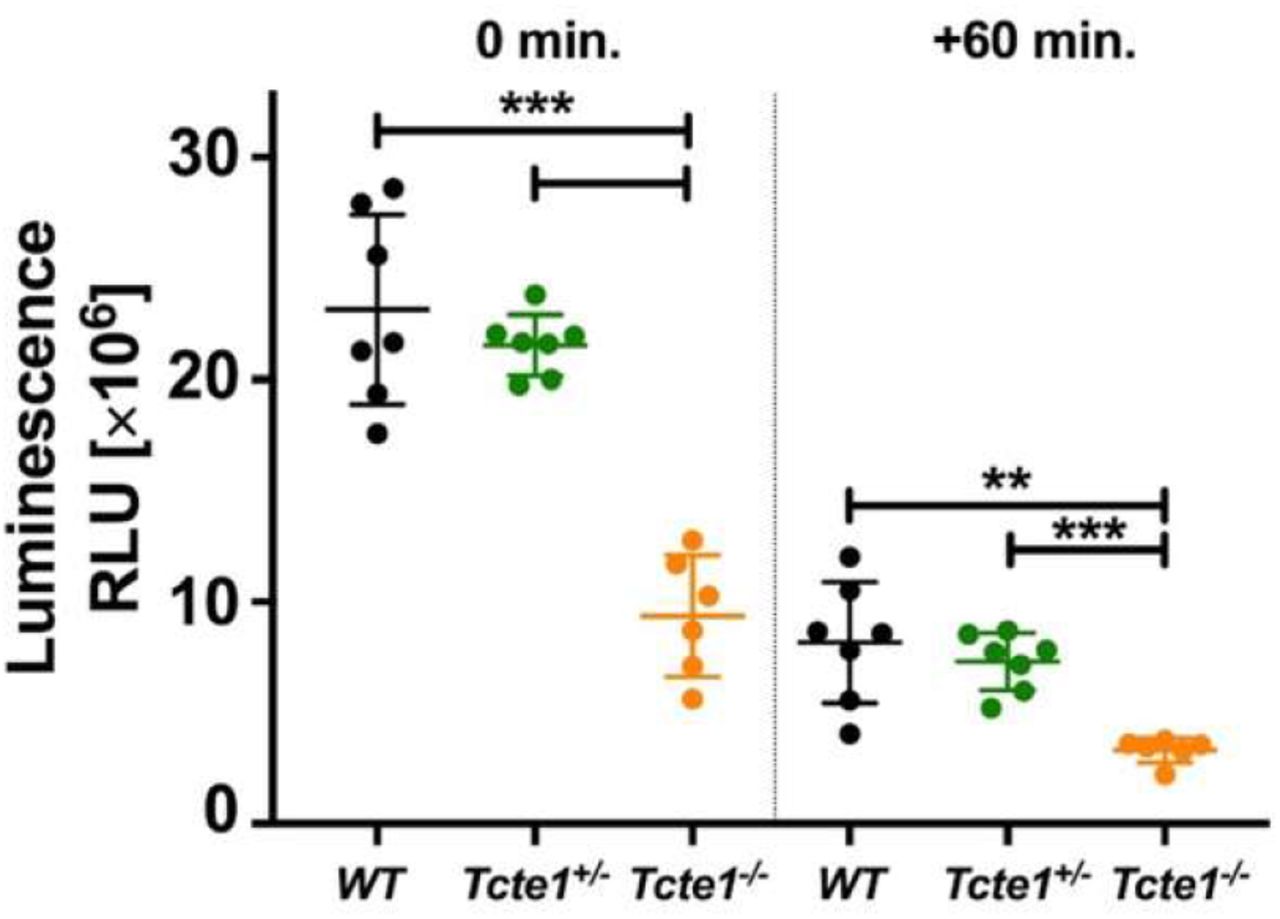
Measurements performed in two time points (0 and +60 min.) revealed that in Tcte1−/− samples the luminescence level was approximately 2.4-fold lower when compared to WT and Tcte1+/− mice (p<0.0001) (Figure 8; Supplementary file 13). There were no differences between WT and Tcte1+/− males. Additionally, the decrease of the luminescent signal after 60 min. was similar in all three groups studied – approximately by 35% (WT: 35.27%, Tcte1+/−: 33.88%, Tcte1−/− 37.76%).

Measurement done in two time points (0 and +60 min.) showed decrease of approximately 35% of luminescence in all three groups. Mean measured values in homozygous group were approximately 2.4-fold lower. Dots – particular animals; long dash – mean value for the group; short dashes – standard deviation; RLU – relative luminescence unit. *** p<0.0001, ** 0.0001 < p < 0.01.
Apoptosis in testicular tissue
Evaluation of the immunofluorescent staining results revealed Casp3-positive signals (indicating apoptosis) in spermatogonia only, in all three mouse genotypes (Figure 9). In each zygosity the numbers of tubules and positively stained spermatogonia (intensity of the signal) were similar suggesting equal levels of apoptosis.

Caspase 3 is one of the apoptotic indicators becuase of its engagement in the induction, transduction, and amplification of intracellular apoptotic signals. Caspase 3 signals were found only on spermatogonia in all three genotypes evaluated (wild type (WT), hetero- (+/−) and homozygous (−/−)). No differences between the level of fluorescent signals were found between evaluated mouse genotypes, indicating similar level of apoptosis. Antibodies: primary (1:100) rabbit anti-Casp3 (Biorbyt, UK); secondary (1:500): goat anti-rabbit-AF594 conjugated (ab150160, Abcam, UK). Fluorescent microscope: Leica DM5500, filters – DAPI, TxR, magnification 630× (with immersion); software: LASX. Bar represents 5 μm. Analysed in 3 independent males per zygosity, on 15-20 tubules each.
Sequencing – human male samples
Potentially associated/important 6 heterozygous SNVs for TCTE1 gene has been found in 15 out of 248 (6.05%) infertile males with drastically decreased semen parameters, ranging from azoospermia to severe oligoasthenozoospermia (Table 1). 3 variants were identified with ultrarare allele frequency (MAF ≤0.0005 in the general population; gnomAD) in three males with NOA: c.862C>T (p.Arg288Ter) predicted as stop-gained mutation leading to non-mediated decay, and thus to protein loss of function with high probability; c.185G>A (p.Arg62His), or c.1048G>A, (p.Gly350Ser), defined as missense ones and probably damaging. Next two SNVs were found as the rare missense ones with MAF <0.005: c.397C>T (p.Arg133Cys) (one NOA case) and c.470A>C (p.Glu157Ala) (four NOA cases), and were defined as probably damaging. The last 1 SNVs (1 NOA, 1 crypto-, and 5 with severe oligoasthenozoospermia) was not found in databases, and thus specified as the novel one (c.374T>G; p.Ile125Arg) with missense prediction (Table 1; Supplementary file 14).
Potentially disease-causing novel and ultrarare heterozygous TCTE1 variants found in patients with disturbed spermatogenesis. Results obtained from screening of n=248 participants. Minor allele frequency data obtained from GnomAD v2.1.1.
SOAs – severe oligoasthenozoospermia; NOA – non obstructive azoospermia; SCO – Sertoli Cell-Only syndrome; Crypto – cryptozoospermia; NMD – non mediated decay; NT – nucleotide; AA – amino acid; SNV – single nucleotide variant; PLoF – Loss of Function probability; rs – Reference SNP accession number; CADD – Combined Annotation Dependent Depletion tool
Protein prediction
To understand the molecular implications of identified mutations on the structure and function of TCTE1, we performed homology modeling experiments using the Phyre2, I-TASSER, and AlphaFold tools [Kelley et al., 2015; Roy et al., 2010; Yang et al., 2015; Yang and Zhang 2015; Tunyasuvunakool et al., 2021]. TCTE1 is predicted to contain at least five tandem leucine-rich repeats (LRR) in its C-terminus [UniProt 2019], while no known three-dimensional protein folds have been assigned to its N-terminus. All homology searches yielded a polypeptide with tandem leucine-rich repeats with nearly 100% confidence (Figure 10A; Supplementary file 15). Modeling the N-terminal region of TCTE1 was more difficult; although both Phyre2 and I-TASSER confidently predicted a largely α-helical secondary structure for the N-terminal half of TCTE1 (Figure 10A; Supplementary file 15), neither tertiary structure was predicted with high confidence; indeed, comparison of the two demonstrates vastly different folds for the N-terminus from the programs. However, the confidence of the AlphaFold-generated model was high for the region just N-terminal to the LRR. Further, based on the predicted aligned error plot for calculated for its models, the confidence of the predicted structure of residues 100-200 was high relative to the position of residues 220-480, providing support for the modeled through-space three-dimensional interactions. That said, we interpreted these models very conservatively.

Secondary structure diagrams and associated confidence schema generated by I-TASSER [46,47,48], Phyre2[45], and AlphaFold [49,50] intensive mode. Positions of amino acids of interest are indicated. Green, predicted unstructured regions; yellow rectangles, predicted α-helices; orange arrows and arrowheads, predicted β“strands. Confidence scale ranges from white to cyan, with the probability of the secondary structure increasing with increasing intensity of cyan; B Cartoon diagram of homology model of human TCTE1 generated by AlphaFold with residues of interest shown as spheres and labeled. N, amino terminus; C, carboxy terminus; C-F Zoomed views of indicated residues and mutations: C Ile-125 resides deep within a hydrophobic pocket, which would be disturbed upon substitution with arginine; D Mutation R133C would impact the hydrogen bond interactions that organize the architecture of the globular domain N-terminal to the tandem leucine-rich repeats; E Mutation of Glu-157 to alanine would destabilize the charge of the surface of this globular region, likely leading to unfolding; F Gly-350 allows for the compact interactions of tandem α-helices of the LRR. Depending on the rotamer it adopted, a serine side chain in this position would sterically clash with residues D321, A324, or V346 in the neighboring helix.
We attempted to hypothesize how mutations of interest could affect the tandem LRRs and putative secondary structure of the N-terminus of the protein. Residue R64 resides in a predicted unstructured region and as such most likely is solvent exposed in the tertiary structure (Figure 10A, B). Mutation of this residue from arginine to histidine could therefore affect the surface potential of this disordered region, making the it less positively charged. Indeed, this mutation would be expected to impact the interactions that the N-terminus of TCTE1 can have with other molecules, including other members of the N-DRC. The interaction of microtubule-associated proteins, including the N-DRC, with binding partners is known to be largely charge-based [Kubo et al., 2017; Drechsler et al., 2019].
Three mutations of interest occur in the globular region just N-terminal to the LRR. The AlphaFold model clearly shows how mutation I125R would affect the structure of this region Figure 10a, c). Residue isoleucine 125 is predicted to be nestled within the hydrophobic core including L97 L109, L112, F91, W131, L197, Y130, and the methyl group of the T121 sidechain. A substitution of positively charged arginine for isoleucine in this position would not be accommodated in this hydrophobic region, likely resulting in local unfolding that could potentially propagate beyond the pocket. Likewise, residue R133 supports the structure of this putative globular domain by forming hydrogen bonds with the backbone of residues 109, 110, and/or 112, thereby possibly contributing to the overall organization of this domain and its positioning relative to the LRR (Figure 10d). Mutation of this residue to a cysteine would remove the hydrogen bonding groups supporting these interactions, likely leading to a local unfolding that could possibly propagate farther. Interestingly, residue E157 is predicted to interact with residues in the LRR, likely stabilizing positively charged residues around it, including several from the linker leading up to the first β-strand of the LRR (Figure 10B, E). Mutation of this residue to an alanine would lead to a tertiary destabilization, most likely affecting the overall structure of the N-terminal region of the LRR and the globular region just N-terminal to it.
Two mutations of interest affect the LRR helices. Mutation R288* results in premature truncation, eliminating most of the LRR region from the final protein product, although it is predicted that this premature termination codon would lead to nonsense mediated decay. Mutation G350S would be expected to result in the destabilization of inter-repeat interactions (Figure 10F), possibly even unfolding the protein from that position on; the lack of a side chain in glycine allows for the close approach of these tandem structural elements, whereas the addition of the serine side chain would abrogate the tight packing of residues, regardless of which rotamer it adopts. This could result in a restructuring its C-terminus and/or deforming the surface of the module, therefore affecting its interactions with its binding partners.
Since the N-DRC is conserved from single-celled eukaryotes to mammals, we took advantage of published cryo-electron tomography studies examining the N-DRC from model organism Chlamydomonas (1). This study was of great interest since it explicitly located the position of the C-terminus of DRC5 with a nanogold particle; importantly, it was consistent with earlier cryo-ET experiments positioning the N-DRC (2). While the resolution of this structure is only 40Å, it was still useful for placing the modeled TCTE1 structure in the context of the entire N-DRC (Supplementary file 15B). Manual docking of the AlphaFold predicted TCTE1 structure is consistent with the cryo-EM tomography data [Gui et al., 2019; Heuser et al., 2009].
DISCUSSION
In this study we have investigated the role of Tcte1 in male infertility using mouse knockout model. Using homo-and heterozygous animals, their reproductive potential has been examined, noting no progeny by homozygous males. While there were no differences in histology of the testes, the size of the gonads varied between the zygosity of animals, followed by two semen phenotypes revealed within the sperm quality evaluation. RNAseq analysis of the mutant testicular tissue demonstrated the influence of Tcte1 mutations on the expression pattern of genes related to mitochondrial cytochrome oxidase activity, apoptosis and spermatogenesis. The ATP level estimate of spermatozoa showed its decrease in homozygous males. Additionally, protein prediction modeling of identified variants in human samples revealed changes in the protein surface charge potential, leading to disruption in helix flexibility or dynamics, and thus, suggesting the disrupted TCTE1 interaction with its binding partners within the axoneme. The major findings concerning observed abnormal sperm tail beating in KO Tcte1 mice were summarized in Figure 11. We have also identified and discussed novel functions of genes with testicular expression significantly disturbed in the Tcte1 knockout. Therefore, this study contains novel and the most complete data concerning TCTE1 role in male infertility, so far.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Summary of findings revealed for abnormal sperm tail beating in mouse knockout model of Tcte1 gene, a component of the N-DRC complex in sperm tail. Three major groups of factors required for proper sperm tail beating were marked with bold colour arrows (green, yellow and blue). Documented connections (solid grey arrows) revealed on the basis of: RNAseq data for testis tissues, protein in silico predictioning for mutated Tcte1 protein, and measurements of ATP level and sperm cell components in mouse spermatozoa. Unresolved possible linkages (cause/effect) marked with question (?) mark.
Phenotypes of homozygotes and heterozygotes
There was only one published study so far, concerning knockout mouse model of Tcte1 [Castaneda et al., 2017], with Authors the infertile homozygous males due to asthenozoospermia derived from limited and circular flagellar beating, and fertile heterozygotes. The study did not find any differences between homozygous and WT males in the testicular histology, followed by similar testis weight, sperm concentration and sperm morphology [Castaneda et al., 2017]. We also have not found any differences in the testicular and epididymal tissues histology. However, the weight of the testes was decreased significantly (∼41% in Tcte1−/−, 7% in Tcte1+/−), followed also by testis size reduction in Tcte1−/− animals (by 22%) (Figure 4B). When comparing semen parameters (Figure 4C-E), we have found enormous decline in sperm concentration in both: Tcte1−/− (3.3-fold) or Tcte1+/− (2.5-fold) males, and double increase in abnormal sperm morphology in Tcte1−/− animals. Similarly, to Castaneda et al. [2017], we have documented the impaired sperm motility representing circular mode of the sperm tail beating. Also, the increased ratio of immotile spermatozoa in homozygous males was documented in our study. We can suggest that these discrepancies may result from various number of animals used for evaluation: 3 males in study of Castaneda et al., [2017] vs. at least 18 per each zygosity in our study, when comparing semen parameters and tissue characteristics (except of sperm morphology evaluation with at least 5 animals used). Probably, also the differences in knockout construction used could be causative for observed variances: reporter-tagged insertion with conditional potential vs. the exon 3 deletion. Thus, we have observed two various phenotypes dependent on the zygosity of the animals: homozygous males manifested oligoasthenoteratozoospermia (OAT) and were infertile, while heterozygous males demonstrated oligozoospermia (O) and remained fertile. Data collected in International Mouse Phenotypic Consortium (IMPC; a repository of all available mouse phenotypes created, followed with the possible linkage to human disease defects; https://www.mousephenotype.org), concern only one Tcte1 mouse study [Castaneda et al., 2017], while we reveal an unmatched phenotypes of decreased sperm count.
We suggest that similar effect – decreased sperm count linked to TCTE1 mutations, could be observed in humans. Our pilot mutation screening of 248 human infertile males with severely decreased sperm count (from oligo-, via crypto-, to azoospermia) revealed 6 heterogeneous SNVs (1 novel, 3 ultrarare, and 2 rare; all predicted as disease causing) with the total frequency of 6.05% (Table 1). We propose that molecular epidemiology studies of infertile patients should reveal the existence of homozygous or a compound heterozygous TCTE1 variants with clear direct downgrading effect on spermatogenesis. However, we claim that also heterozygous variants can be causative, similarly to SPINK2 which deficiency has been documented as responsible for oligoasthenozoospermia in heterozygous mice vs. azoospermia in homozygous animals, followed by the azoospermia in human homozygous brothers vs. their heterozygous father with oligozoospermia [Kherraf et al., 2017]. It can be explained by the probable haploinsufficiency occurrence, when the single copy of the WT allele in heterozygous combination of which a variant allele is insufficient to produce the WT phenotype. Our observations, followed by the SPINK2 data and data related to infertility (i.e.: P1/P2 protamines, Pde8b, Trim28, Tcfl5), and to the other diseases (i.e.: PIKFYVE in cataract, NFIA, SCN1A in neurological or epilepsy disturbances, KLF1 in hemoglobin persistenece, NR5A2 in pancreatic cancer), seem to confirm that the severity of the phenotype depends on the gene expression level [Jodar and Oliva, 2014; Mei et al., 2022; Ogura et al., 2022; Heshusius et al., 2022; Valassina et al., 2022; Sandhu et al., 2021; Leal et al., 2021; Tan et al., 2020; Xu et al., 2022]. We have documented variable phenotypes for a novel TCTE1 variant c.374T>G (p.Ile125Arg) that has been documented in seven males, including: azoospermia, crypto-, and severe oligoasthenozoospermics. Thus, it seems to suggest the reasonable candidacy of haploinsufficiency as the mechanism responsible for observed phenotypic spectra also in case of the TCTE1 gene.
Kallikreins, Aquaporin 2
Our pathway analysis revealed a network including two genes with changed expression level in testis: Igf2 (2-fold increased) and Klk1b22 (8-fold decreased), supported by an interesting set of proteins linked to apoptosis and cellular processing (Figure 5; Figure 6; Supplementary file 11; Supplementary file 12). We can suggest that observed Igf2 overexpression in our study can be one of the associated factors leading to decreased sperm count and testis parameters in evaluated KO mice model because of its known relation to spermatogenesis and testis development [Matsuzaki et al., 2020; Cannarella et al., 2020; Neirijnck et al., 2019; Damaschke et al., 2017]. Documented reductions can be supported by the observed constrained expression of Klk1b22 – kallikrein mostly known from its neuronal activity; however, it cannot be excluded that we have revealed additional role of this protein, similar to the other members of the kallikrein family – role in semen liquefaction and degradation of extracellular matrix proteins in the interstitial area surrounding Leydig cells of the adult mouse testes (Klk1b27, Klk1b21, Klk1b24) [Michaelis et al. 2017; Marques et al., 2016, Du Plessis et al., 2013; Fink et al., 2007]. Notably, Klk1b22 expression is significantly enriched in testis. While expression of all four kallikreins was decreased 2.5-8-fold in our Tcte1 KO model, indicating their participation in the reduction of sperm parameters measured (Fig.5).
We have also documented 2-fold decrease of expression of Aqp2 (Aquaporin 2), involved in fluid resorption and water movement within the duct system, with known role in spermatogenesis [Klein et al., 2013; Ramli et al., 2019]. It was documented that in stallions Aqp2 is being expressed in Leydig cells, round and elongated spermatids [Klein et al., 2013]. Also, this protein works within a network of molecules that are linked to cell membrane trafficking (i.e., Rab11a, Myo5b), followed by activity of protein kinases (Prkacb, Prkaca) that are known as regulators of cell proliferation, chromatin condensation/decondensation, and nuclear envelope disassembly/reassembly, and all is supported by the ATP metabolism (Hspa8) [Supplementary file 11] [Markou et al., 2022; Wagner et al., 2022]. Considering this evidence, we suggest Aqp2 play an important role in reduction of the spermatids volume during spermatogenesis and in proper sperm morphology assessement and observed morphological differences between Tcte1 mice zygosities in our study.
Disruptions in sperm cell structure
TCTE1 protein contains five leucine rich repeats (LRRs) that are known to be directly involved in receptor-mediated signaling on the structural basis for interaction between proteins [Ng et al., 2011; Kobe and Kajava 2001]. It was documented that variants in genes encoding LRRs are commonly implicated in the pathogenesis of more than 60 human diseases [Ng et al., 2011; Matsushima et al., 2019]. LRR mutations mostly occur within regions that are protective for the hydrophobic core of the LRR domains. Thus, mutations in LRRs may affect the charge of the surface potential, leading to conformational changes of the protein or induction of protein cumulation. Our prediction model of potential changes in TCTE1 protein resulting from mutations found in human samples seems to point out the prospective impact into the interactions of the TCTE1 with other molecules (binding partners) known to be largely charge-base, including N-DRC elements or microtubule-associated proteins [Kubo et al., 2017; Drechsler et al., 2019]. In the case of TCTE1 protein observed amino acid substitutions promote the abnormal aggregation of TCTE1 protein, and decrease its structural stability, then probably leading to the observed disturbances. Beating motion of flagella is maintained by electrostatic cross-bridge between negatively charged tubulins and positively charged N-DRC [Kubo et al., 2017]. Thus, changes in the charge potential of TCTE1 surface, which is one of the N-DRC components, seem to provide modified (reduced) flagellar beating, similarly as in case of mutations in other N-DRC element – DRC4 protein, of which changes in charge were essential for proper flagellar motility [Kubo et al., 2017; Lewis et al., 2016]. Also, the inflexibility of the midpiece observed in spermatozoa of Tcte1 knockout mice seems to be a consequence of changed structure of LRRs in Tcte1 protein.
It is also known that the lengths of the sperm midpiece and tail may be crucial in determination of sperm swimming velocity, followed then by decreased fertilization rate [Firman and Simmons 2010; Gu et al., 2019]. Available data showed that sperm with shorter midpiece may swim slower and were observed in asthenozoospermic cases [Firman and Simmons 2010; Gu et al., 2019; Anderson et al., 2002]. The meta-analysis of sperm shape features according to sperm competition theory has also shown, that in majority of vertebrates the longer sperm cell (including midpiece or flagellum) has been attributed to faster swimming, followed by the statement that the sperm size may be increased when sperm number is lowered [Lüpold et al., 2020]. In our study we have documented longer sperm tails and midpieces in Tcte1−/− animals, that in fact were infertile. Thus, we can suggest that: (i) the observed longer sperm tails and midpieces may be co-responsible for aberrant motility observed, and (ii) the documented longer sperm tails and midpieces may be an indirect result of constrained sperm count observed, because of some sort of compensation for aberrant motility. However, this issue remains only a suggestion because of a restricted literature data relative to similar observations. Bennison et al. [2016] revealed than in a passerine bird the sperm tail velocity was correlated to total sperm length, but only to some point of the length value at which the velocity started to be decreased. On the other hand, Gu et al. [2019] have shown that in mammalian species there were some changes in the axonemal slopes when the flagellar length increases. The results obtained within our study, and supported by other research concerning sperm tail beating pathways, should be a good start point for creation of further mathematical computations for better understanding of the asthenozoospermia.
Constrained energy metabolism
The flagellar motility is determined by coordinated activity of asymmetrically distributed dyneins in the axoneme. The driving factor for this activity is the hydrolysis of ATP released from OXPHOS system (Mitochondrial Oxidative Phosphorylation System) [Tourmente et al., 2015; Ford 2006]. ATP is being produced in mitochondria placed within sperm midpiece. In our study ATP level measurements revealed its decreased level in sperm cells of Tcte1−/− mice, what can result in faulty dynein functioning, leading to asthenozoospermia via abnormal sperm tail beating. We have documented that Tcte1 KO resulted in significant changes in expression level of testicular genes involved in mitochondrial energy transport. That leads to the conclusion that homozygous Tcte1 mutation results not only in inflexible midpiece leading to asymmetric circular motility, as presented in this study and by others [Fung et al., 2018], and then to asthenozoospermia, but also to aberrant functioning of mitochondrial OXPHOS machinery leading to disruption of ATP production. Thus, the sperm tail has also insufficient power for beating because of the decreased ATP content. Castaneda et al. suggested that the reasons of the observed ATP decrease may result from disturbances in a glycolytic pathway [Castaneda et al., 2017]. It is already known, that glycolytic energy is enough for spermatozoa to survive [Nascimento et al., 2008; Zhang et al., 2022]. However, for sperm motility and then fertilization, the OXPHOS energy production is required [Tourmente et al., 2015; Ford 2006; Zhang et al., 2022]. In our study, RNAseq results clearly revealed the influence of Tcte1−/− mutation on genes involved directly in electron transport chain, such as: mt-Co2 and mt-Co3, crucial components of the respiratory chain complex IV, one of the OXPHOS elements. Observed reduced expression seems to clearly evidence the decreased ATP production via disturbed electron chain transportation, and then revealing the sperm motility reduction observed in Tcte1−/− males, leading to observed asthenozoospermia. Detailed STRING analysis revealed definite interactions between both listed proteins identified in our study, and a list of other mitochondrial ones (mt-Co1, mt-Cytb, Cox5a, Cox6a, Cox7c, mt-Atp6; Supplementary file 11; Supplementary file 12) clearly related to proper processing of cytochrome energetic activity [Brischigliaro and Zeviani 2021; Ganetzky et al., 2019]. Overexpression or mutations in COX1, COX2, and COX3 were previously observed in infertile men with varicocele and OAT, or diabetes [Heidari et al., 2016; Mostafa et al., 2016; Perrotta et al., 2012]. Additionally, it was shown that system of COX2 and prostaglandins plays key role in development of male gonad, spermatogenesis and steroidogenesis, especially in functioning of Sertoli and Leydig cells, and also was found to be significant in the background basis of idiopathic infertility status [Frungieri et al., 2015]. It seems that in our study the reason of observed disturbed motility, represented as circular movement of spermatozoa, supported by increased ratio of immotile sperm cells in Tcte−/− mice, leading to asthenozoospermia, resulted from two factors: (i) inflexibility of sperm midpiece resulted from changed protein charge level in the Tcte1 protein, followed by elevated lengths of sperm tail and midpiece, and (ii) decreased amount of ATP produced, that may be an outcome of inability to get sufficient energy for proper sperm motility.
Protection and/or compensation?
We have also documented an important role of the Fetub gene for the first time in the male testis. Besides of its known role in female fertility [Karmilin et al., 2019; Bansho et al., 2017], the latest study pay attention also in the context of prostate cancer is a detailed way [Zhan et al., 2020]. It was found that overexpression of FETUB increases the apoptosis of prostate cancer cells, followed by the inhibition of their proliferation, migration and invasion, and also slower growth of tumor when compared to controls [Zhan et al., 2020]. Thus, in the overexpression mode, the Fetub plays protective role for the prostate. It seems to explain results obtained in our study, where the Fetub expression was increased almost 5-fold in Tcte1−/− mice, but no tumor signs have been observed in the mouse testis at the tissue level. Zhan et al. [2020] have found the connection between Fetub overexpression, and induction of Bax (Bcl-2-associated X protein) and capases, resulting in promoting the apoptosis. It is known that the Bax/Bcl-2 induction/inhibition is directly related to the release of cytochrome c (Cyt c) in the caspase-dependent signaling [Dadsena et al., 2021; Samaiya et al., 2021; Li et al., 2021]. And cytochrome c is one of the two electron carriers in electron transport chain responsible for transport directly to the respiratory chain complex IV [Yin and O’Neil 2021; Alvarez-Paggi et al., 2017]. Due to Cyt c release, complex IV is essential for apoptosis and cell fate [Samaiya et al., 2021; Li et al., 2021]. Following of all of those facts, our results seem to reveal the connection between Fetub and diminished mitochondrial chain IV activity in mice testis, what have not been reported so far. It can be also supported by the fact of the known relation between FETUB and the inactivation of the PI3K/AKT signaling pathway (key player in regulation of cell proliferation and apoptosis in cancerogenesis), in which the Bax/Bcl-2 are involved [Zhan et al., 2020].
The novel testis-protective role in our study can be also credited to three other genes with increased expression level in testicular tissue: Hoxd10 (3-fold increase), Slc45a3 (2.2-fold), and Ube2l (2-fold). Overexpression/activation of HOXD10 has been already documented as inhibitor of tumorigenesis [Jonkers et al., 2020; Mo et al., 2017; Zhang et al., 2019]. Expression of Slc45a3, known also as ‘prostein’, has been documented as weaker in cancer tissues [Hernández-Llodrà et al. 2017; Pin et al., 2017; Perner et al., 2013]. Elevated expression level of Ube2l is known to be connected with the degradation/suppression of tumor cells [Li et al., 2018]. On the other hand, the 12-fold increased gene expression level for Usp39 (ubiquitin specific peptidase 39) – an oncogenic splicing factor which expression is positively linked with cancerogenesis’ promotion in prostate or ovaries, while its silencing induces apoptosis and cell cycle arrest, has been noticed [Huang et al., 2016; Yuan et al., 2021; Zhao et al., 2016].
Taken together data obtained for genes related to cancerogenesis and/or apoptosis described above, we can hypothesize that we observe an interesting novel network of interactions between two groups of genes that are reciprocally neutralizing potential influence onto the testis tissue. Mainly, in the first group we have genes with suggested novel anti-cancer role in the testis (Fetub, Hoxd10, Slc45a3, Ube2l6), which increased expression level inhibits the cancer cell development, supported by protective role of Mt3; while in the second corner of the ring the Usp39 gene with elevated expression level (the most from all the observed changes) should lead to reveal some tumor-like changes within the testis tissue. Following the fact, that both: histological evaluation, as well as caspase 3 staining (Fig.9), have shown no differences between hetero- and homozygous mutant mice, our hypothesis seems to be justifiable. On the other hand, there is a prediction that many of the genes (especially so-called cancer-testis associated antigens, CTA) related to spermatogenesis may function in tumorigenesis [Aurriere et al., 2021; Yang et al., 2020; Babatunde et al., 2017; Wang et al., 2016; Bode et al., 2014; Cheng et al., 2011; Nagirnaja et al., 2018]. Due to known similarities between oncogenic and spermatogenic processes, especially in the energy metabolism, we can carefully suggest/hypothesize that in our KO model the genes with changed expression profile found, known as highly involved in tumorigenesis, may also be important in spermatogenesis. Obviously, there is a need for further studies in this direction to confirm these suggestions or to reveal the other (still unknown) mechanisms.
Conclusions
We have documented that TCTE1 gene is the next one that should be added to the ‘male infertility list’ of molecular background because of its crucial role in spermatogenesis and proper sperm functioning. The mutations in mouse Tcte1 knockout model revealed two phenotypes, depending from zygosity with infertile oligoasthenoteratozoospermic homozygotes, and fertile oligozoospermic heterozygotic males, suggesting haploinsufficiency mechanism involvement. All collected data pointing that the Tcte1 protein is functioning in spermatogenesis and show how its mutations add the level of complexity to the clinical manifestation of decreased semen parameters. The disrupted energy machinery of the spermatozoa, followed by newly suggested roles of genes related to cancerogenesis and/or apoptosis (here in the context of their neutralizing/compensation effect on the testicular tissue), seem to underline the complex and devastating effect of the single-gene mutation, on the variety of testicular molecular networks, leading to reproductive failure.
Data Availability
All data generated or analyzed during this study are included in this published article and its supplemental information files. Supplementary files are deposited in Zenodo at https://doi.org/10.5281/zenodo.7330637 RNAseq data are available in GEO database (https://www.ncbi.nlm.nih.gov/geo/) under the accession number: GSE207805. The results described in the publication are based on whole-genome or exome sequencing data which includes sensitive information in the form of patient specific germline variants. Information regarding such variants must not be shared publicly following the European Union legislation described in the following document: https://publications.jrc.ec.europa.eu/repository/bitstream/JRC113479/policy_report_-_review_of_eu_national_legislation_on_genomics_-_with_identifiers_1.pdf, therefore the access to raw data that support findings of this study are available from the corresponding author upon reasonable request.
Authors’ contributions
MO design of the study, manuscript drafting and editing, coordination of the tasks, data interpretation, gene selection, preparation of human blood and semen samples, mice: mating, genotyping, gross sections and preparation of biological samples, RNAseq analysis, immunofluorescence, Sanger sequencing, ATP measurements; AM gene selection, data interpretation, genotyping, RNAseq analysis, collection and preparation of human testis samples, WGS and Sanger sequencing, manuscript editing; TS bioinformatic analysis of RNAseq and WGS results; NP WES of azoospermia samples (incl. bioinformatics); AB protein prediction modeling and its description; SB preparation of: DNA for sequencing, RNA for RNAseq, screening of databases for RNAseq results; MKa collection of human blood and semen samples, semen analyses of human and mouse samples, detailed morphology of mouse spermatozoa; HJ histopathology of mouse tissue sections; PJ recruitment of patients and their medical history; ANY gene selection, interpretation of azoospermia WES results, data interpretation; MKu data interpretation, gene selection, recruitment of patients and their medical history, correction of the manuscript, funds collection and supervision.
All authors critically reviewed and approved the final version of the manuscript.
LARGE SCALE DATA
All data generated or analyzed during this study are included in this published article and its supplemental information files.
RNAseq data are available in GEO database (https://www.ncbi.nlm.nih.gov/geo/) under the accession number: GSE207805.
The results described in the publication are based on whole-genome or exome sequencing data which includes sensitive information in the form of patient specific germline variants. Information regarding such variants must not be shared publicly following the European Union legislation described in the following document: https://publications.jrc.ec.europa.eu/repository/bitstream/JRC113479/policy_report_-_review_of_eu_national_legislation_on_genomics_-_with_identifiers_1.pdf, therefore the access to raw data that support findings of this study are available from the corresponding author upon reasonable request.
LIMITATIONS, REASONS FOR CAUTION
In the study, the in vitro fertilization ability of homozygous male mice was not checked.
WIDER IMPLICATIONS OF THE FINDINGS
This study contains novel and the most complete data concerning TCTE1 role in male infertility. TCTE1 gene is the next one that should be added to the ‘male infertility list’ of molecular background because of its crucial role in spermatogenesis and proper sperm functioning.
STUDY FUNDING/COMPETING INTEREST(S)
This work was supported by National Science Centre in Poland, grants no.: 2015/17/B/NZ2/01157 and 2020/37/B/NZ5/00549 (to MK), 2017/26/D/NZ5/00789 (to AM), and HD096723, GM127569-03, NIH SAP #4100085736 PA DoH (to AY). The authors declare that there is no conflict of interest that could be perceived as prejudicing the impartiality of the research reported.
Supplementary files legend
Supplementary File 1: Vectors used for knockout creation of Tcte1 gene.
Supplementary File 2: Real time PCR conditions used for determination of relative expression level of Tcte1 gene in mouse knockout animals.
Supplementary File 3: Reproductive potential of KO Tcte1 mice. Mating combinations and their results including: time of breeding, number of litters and pups, number of male and female pups, and genotypes observed in pups of each combination.
Supplementary File 4: Genotyping protocol for Tcte1 knockout model.
Supplementary File 5: Characteristics of antibodies used for N-DRC complex staining in mouse sperm. Human antigen sequence identity between human and mouse orthologs was also shown (blast comparison; information from Biorbyt producer).
Supplementary File 6: Schematic models of exon 3 deleted in KO mice. A Tcte1 protein exon 3 (deleted in KO) prediction of 5 top models (software: I-TASSER). C-score is the confidence score of the prediction. C-score ranges [0-1], where a higher score indicates a more reliable prediction; TM-score is a measure of global structural similarity between query and template protein., RMSD - root-mean-square deviation of the average distance between the atoms. B The deletion sequence used for modeling, including exon 3 (yellow).
Supplementary File 7: An example of circular type movement pathway of spermatozoa from Tcte1−/− male mice.
Supplementary File 8: An example of sperm cells motility in Tcte1+/− male mice.
Supplementary File 9: An example of sperm cells motility in WT male mice.
Supplementary File 10: Detailed parameters of mouse sperm morphology. Statistically significant values: p < 0.05; ns – statistically not significant (p ≥ 0.05). For samples with normal distribution (positive Shapiro-Wilk normality test), t-test with Welch’s correction was applied; for samples without normal distribution (midpiece and tail defects), Kolmogorov-Smirnoff test was applied (GraphPad Prism v. 7.0e). n=200 spermatozoa were analyzed per each mouse; for WT and Tcte1−/− n=5 animals were evaluated, while for Tcte1-/+ the number of animals was n=7. Spermatozoa stained with the classic Papanicolau staining procedure. Bright field microscope: Leica DM5500, magnification 630x (with immersion), software: CytoVision.
Supplementary File 11: STRING analyzis data for each gene with changed expression pattern in Tcte1 KO mouse model.
Supplementary File 12: STRING analyzis of potential interactions between proteins with observed major differences in gene expression level in mouse testis of the homozygous Tcte1 mice (Usp39, Fetub, Klk1b22). According to genes revealed in RNAseq analysis (marked with arrows), two groups of interactions were formed for genes with highest expression changes (red arrows); (STRING database, 11th March 2021).
Supplementary File 13: ATP measurement data obtained for KO Tcte1 male mice. wt = wild type, het = heterozygous, ho - homozygous. RLU = relative luminescence unit.
Supplementary File 14: Examples of potentially disease-causing rare heterozygous TCTE1 variants found in patients with disturbed spermatogenesis. Results obtained from screening of n=248 participants. A novel variant (Sanger sequencing); B ultra rare variants (WES).
Supplementary File 15: The C-terminal region of TCTE1 was consistently modeled and fits well in published maps. A Superposition of the cartoon representation of three models of human TCTE1 generated by Phyre2 [45] intensive (green) or fast mode (grey), I-TASSER [46,47,48] (yellow), and AlphaFold [49,50] (cyan). N- and C-termini are labeled. B. Manual docking of the model of full-length TCTE1 generated by AlphaFold in maps calculated from a cryo-electron tomography experiment performed on C. reinhardtii using nanogold to locate the C-terminus of DRC5 (EMD-20821) [23].
ACKNOWLEDGEMENTS
We are grateful to Oliwia Kordyl, MSc, and Katarzyna Kurek, MSc (IHG PAS, Poznan) for their technical help in preparation of the samples for sequencing, genotyping, and immunofluorescence staining.
Footnotes
other authors’ emails: Marta Olszewska, PhD marta.olszewska{at}igcz.poznan.pl, Agnieszka Malcher, PhD agnieszka.malcher{at}igcz.poznan.pl, Tomasz Stokowy, PhD tomasz.stokowy{at}uib.no, Nijole Pollock, BSc Nijole.pollock{at}gmail.com, Andrea J. Berman, PhD AJB190{at}pitt.edu, Sylwia Budkiewicz, MSc sylwia.budkiewicz{at}wp.pl, Marzena Kamieniczna, PhD marzena.kamieniczna{at}igcz.poznan.pl, Hanna Jackowiak, PhD hanna.jackowiak{at}up.poznan.pl, Piotr Jedrzejczak, PhD, MD piotrjedrzejczak{at}gmail.com, Alexander N. Yatsenko, PhD, MD yatsenkoan{at}mwri.magee.edu
REFERENCES




