
Abstract
Background Mobile colistin resistance (mcr) genes modify Lipid A molecules of the lipopolysaccharide, changing the overall charge of the outer membrane.
Methods A systematic review of all studies published between January 2015 to July 2021 was performed. Included articles described mcr genes in the context of their genetic environment, fitness cost, crystal structure, their enzymatic activity and the risk factors associated with the acquisition of mcr. Studies describing the epidemiology of mcr genes and novel therapeutics were included.
Results and Discussion Ten mcr genes have been described to date within eleven Enterobacteriaceae species, with Escherichia coli, Klebsiella pneumoniae, and Salmonella species being the most predominant. They are present worldwide in 72 countries, with human specimens currently having the highest incidence. This is due to the wide dissemination of mcr in livestock animals, meat, manure, the environment, and wastewater samples, increasing the risk of transmission via foodborne, zoonotic, and vector-borne routes to humans. The stability and spread of mcr genes were mediated by mobile genetic elements such as the IncHI2 conjugative plasmid, which is associated with multiple mcr-variants and other antibiotic resistance genes. The cost of acquiring mcr is reduced by compensatory adaptation mechanisms. MCR proteins are well conserved via structurally. Hence, MCR-1 inhibitors and therapeutics should be applicable to all MCR proteins.
Conclusion Mcr genes have spread from animals into the clinical setting, threatening public health. Combination therapies are a promising option for managing and treating colistin-resistant Enterobacteriaceae isolates whilst reducing the toxic effects of colistin.
Importance/Highlights
Mcr genes are associated with mobile genetic elements that are facilitating its global dissemination.
Using colistin as a growth promoter increases the risk of acquiring mcr-positive Enterobacteriaceae in food-producing animals.
There is a higher incidence of mcr genes in humans than animals.
MCR proteins are phosphoethanolamine (PEtN) transferases that mediate the transfer of PEtN from its primary phosphatidylethanolamine to lipid A.
Multiple compounds can synergistically restore colistin’s activity, reducing its dosage and toxicity.
CRISPR-Cas9, endolysins-engineered enzymes, and antimicrobial peptides are promising therapies for colistin resistance
Tweet: “Within bacteria, mobile colistin resistance genes have spread widely, with a high incidence in human samples. It’s critical to comprehend the genetic tools facilitating this spread and their mechanisms of action. These are detailed in this paper, as are new therapeutic strategies for managing these resistance genes.”
Introduction
Colistin was first introduced into clinical practice in the 1950s 1. It was derived from Bacillus polymyxa and belongs to polymyxins 2, a family of cationic polypeptide antibiotics with broad-spectrum antimicrobial activity 3. Colistin has a bactericidal effect on Gram-negative bacteria and thus is used for treating Gram-negative bacterial infections 1, 2. Cationic polypeptides have a high electrostatic attraction to the anionic lipopolysaccharide (LPS) located on the outer membrane of Gram-negative bacteria. There, it displaces the magnesium and calcium divalent cations (Mg2+ and Ca2+), which stabilise the LPS molecules 1, 2. This results in the disruption of the cell’s permeability, leading to cell death. However, due to the adverse side effects such as nephrotoxicity and neurotoxicity seen during colistin therapy 2, in the early 1980s, it was removed from human use and administered to food-producing animals as a growth promoter and therapeutics 1, 4. With the increasing resistance caused by Gram-negative pathogens such as carbapenem-resistant Enterobacteriaceae that threatens global public health 5, colistin was recently reintroduced as a last-line treatment option 3, 4, 6.
Colistin resistance was largely associated with chromosomal-encoded mechanisms that involved two component systems (TCSs) such as pmrAB and phoPQ, and mutation(s) in the mgrB regulator in Klebsiella pneumoniae 7–9. These chromosomal mutations mediated colistin resistance by modifying LPS, changing LPS’s overall charge, and reducing the affinity of polymyxins to the outer membrane 7–9. The types of modifications seen include the addition of phosphoethanolamine (PEtN) and 4-amino-4-deoxy-L-arabinose (Ara4N) to the 1-phosphate or 4-phosphate groups of Lipid A, respectively 10. The PEtN modification is associated with the pmrAB TCSs and the Ara4N modification with the phoPQ TCS alongside the mgrB regulator 10.
Plasmid-mediated colistin resistance (mcr) gene was first identified in both animals and humans by Liu et al. (2016) 11 from an Enterobacteriaceae. mcr-1 increased colistin resistance and encoded a PEtN transferase enzyme that added PEtN to Lipid A 11 at the 4’-phosphate group 12, 13. The MCR-1 PEtN transferase enzyme had similar structural properties to EptC and LptA PEtN transferases from Campylobacter jejuni and Neisseria meningitidis, respectively 14, 15. The lipid A modification is identifiable with a matrix-assisted laser desorption/ionization-time of flight (MALDI-TOF) mass spectrometry (MS) assay with an additional m/z= 1920.5 peak observed in mcr-producing isolates, representing the modified Lipid A molecule 16. This activity is seen across the different mcr variants: mcr-2 to mcr-10.
After the discovery of mcr-1, other novel mcr variants i.e., mcr-2 to mcr-10, which are widely distributed within Enterobacteriaceae, have been reported globally. The identification of the mcr-1 gene and its variants, mcr-2 to mcr-10, is largely mediated by PCR screening and whole genome sequencing (WGS) tools, which are also used in mcr surveillance programmes 11, 17–21. Each mcr variant is mostly located on conjugative plasmids, associated with mobile genetic elements (MGEs), and mediates colistin resistance through PEtN transferase activity. mcr are widely distributed within Enterobacteriaceae, including Escherichia coli, K. pneumoniae, Salmonella species, and Enterobacter species, 3, 11, 22, 23 and have also been reported within other Gram-negatives such as Pseudomonas aeruginosa and Acinetobacter species 24–26.
Initially, food-producing animals were the reservoir of mcr genes due to the high usage of colistin in livestock 11, 27, 28. Farmers’ consumption of livestock and/or contact with livestock or their faeces was found to be a risk factor for infection with an mcr-producing isolate 28, 29. Global screening of mcr-1 genes in livestock found increased number of mcr-producing isolates, resulting in a ban of colistin in food-producing animals for both growth promotion and treatment of bacterial infections 30, 31. The World Health Organisation (WHO), thereafter, listed colistin as part of the critically important antimicrobials for human medicine. This was to help preserve the effectiveness of colistin for clinical use and to minimize the transmission of mcr genes from animals, livestock, and the environment to humans 30, 31.
Purpose of review
This review provides a map of the dissemination of mcr-1 and its variants. It further evaluates the genomic content of each mcr variant, identifying the possible progenitor and the mobile elements that each is associated with. We largely look at the mcr-1 gene, its structure and function, which enables it to mediate colistin resistance, the fitness cost imposed by mcr-1 expression, and the risk factors enabling the dissemination of mcr genes. The review further summarizes the possible treatment options for mcr-producing colistin-resistant isolates.
Herein, we highlight the epidemiology and evolution of mcr genes over the last 6 years by providing insight into the genomic content of each variant, identifying MGEs that aid in its dissemination, their global distribution, the crystal structure, and enzymatic activity of MCR proteins, and promising emerging therapeutics that could manage mcr-positive Enterobacteriaceae infections.
Methods
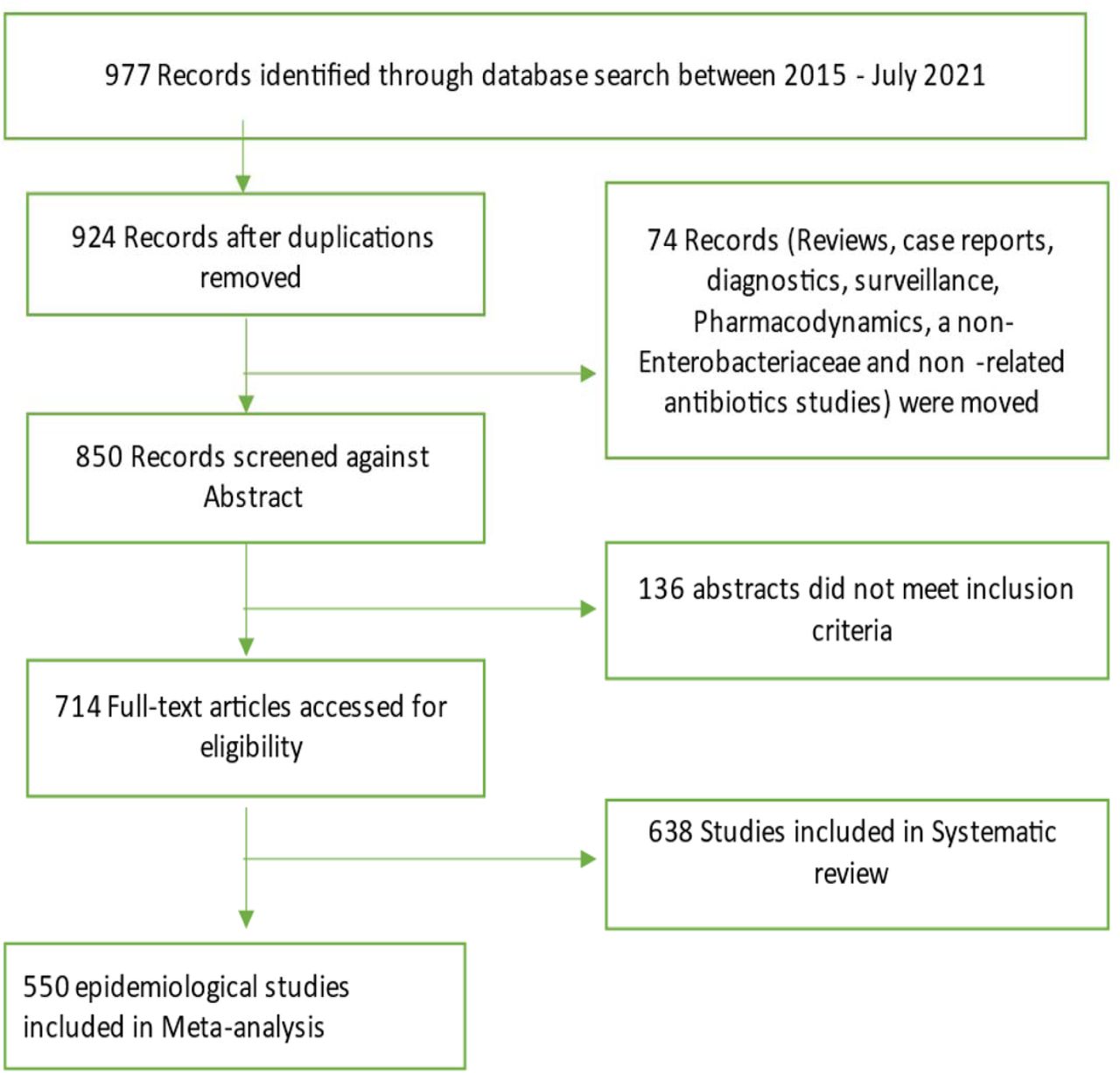
A comprehensive literature search was performed using PubMed. Journal articles published in English within the last six years (January 2016 to July 2021) were retrieved and screened using the following keywords: “colistin resistan*” or “polymyxin resistan*” and “mcr*” and “Enterobacteriaceae”. The search was focused on journal articles that discussed the crystal structure of MCR enzymes and their enzymatic activity, the genetic environment and genetic support (MGEs) of mcr genes, as well as their molecular epidemiology, risk factors and management. However, studies that involved reviews, books and documents, case reports, case studies were excluded. The inclusion and exclusion protocol used in this review is seen in Figure 1. The data that was extracted from the included studies is found in Table S1.

Flow diagram of literature search strategy and methodology. Out of 977 identified records between 2015 and 2021, a total of 638 studies were included in the systematic review whilst 550 of this were used for the epidemiological analyses.
Results
Characteristics of included studies
The study included 693 articles describing the epidemiology of mcr genes, the crystal structure and enzymatic activity of MCR proteins, the genomic environment of each mcr gene, the risk factors associated with the acquisition of mcr-positive Enterobacteriaceae isolates and the novel therapeutic options that can be explored for managing mcr-positive Enterobacteriaceae-related infections. A total of 429 articles were included in the epidemiological data and were used for the statistical analysis. The following data was extracted from the epidemiology articles: country, study year, specimen type and source, bacterial species, clone/MLST, mcr-type, mobile genetic elements (MGEs) and antibiotic resistance genes (ARGs) (Table S1).
Enterobacteriaceae species distribution
mcr genes are well disseminated within Enterobacteriaceae, being frequently identified within E. coli, K. pneumoniae and Salmonella species, in that order. Other Enterobacteriaceae species that have been identified with mcr genes include Cronobacter species, Citrobacter species, Kluyvera species, Leclercia species, Raoultella ornithinolytica, and Shigella species (Figure 2A).

The sources and types of Enterobacteriaceae species identified to be harbouring mcr genes. A) Sources reported to harbour mcr-positive Enterobacteriaceae isolates were mainly animals, humans, water, food, and environmental samples. B) Distribution of mcr-producing Enterobacteriaceae species per sample source. C) The number of reports for each mcr-positive Enterobacteriaceae species.
Initially, mcr genes were commonly identified in animals. However, after evaluating the number of reports collected from 2016 to 2021, the most common source of mcr-positive Enterobacteriaceae isolates was human specimens (Figure 2a). In human specimens, mcr genes are frequently isolated from Enterobacter sp., K. pneumoniae, Leclercia sp., and Shigella sp (Figure 2b). E. coli is the most common mcr-positive isolate and is usually isolated from animal specimens (Figure 2c); this is also seen with R. ornithinolytica, Salmonella sp., and S. Typhimurium (Figure 2b). Other sources identified include food and the environment, which are mostly made up of wastewater samples. In the environment, mcr-positive isolates comprised only three species: E. coli, K. pneumoniae, and Kluyvera sp (Table S2). Lastly, mcr-positive species have been also isolated from food animal and vegetable samples, being isolated from mostly packaged meat and vegetables. These species include: Cronobacter sp., Enterobacter sp., E. coli, K. pneumoniae, Salmonella, and S. Typhimurium.
Mcr-1 and its variants are the most common and well disseminated of all mcr variants (Figure 3a). This was later shown to be because of multiple factors such as fitness cost and MGEs. Mcr-1 has spread globally and has been identified in 69 countries, with mcr-1 being the most prevalent mcr variant in most countries (Figure 3c). Although the other variants have spread globally, they have been identified in low numbers compared to mcr-1 (Figure 3a). This includes mcr-3, which has been identified in fifteen countries, with Colombia and Thailand having the highest counts. Mcr-3 variants (mcr-3.1, mcr-3.5, etc) have been identified in an additional three countries, and in a total of eight countries. This brings the total count of countries with mcr-3 genes to eighteen (Table S3, Figure 3c). Other well disseminated mcr variants in low numbers includes mcr-5 and mcr-9, identified in ten countries. mcr-8 has been reported in seven countries, mcr-4 in five countries, mcr-7 and mcr-10 in two countries, and mcr-6 in Thailand only.

Global distribution and total count of mcr genes. A) Total number of mcr genes reported globally. B) The distribution of mcr genes per country. C) Global map showing the geographical distribution of mcr genes.
Amongst the 70 countries identified with mcr genes, countries such as Thailand harboured six mcr-variants (mcr-1, mcr-3.mcr-6, mcr-7, mcr-8 and mcr-9) and China harboured eight variants (mcr-1, mcr-3, mcr-4, mcr-5, mcr-7, mcr-8, mcr-9 and mcr-10). Other countries such as the USA, Turkey, Spain, Nigeria, Korea, Japan, Italy, France, England, Czech Republic, Cambodia, Brazil, Belgium, and Bangladesh harboured three to four mcr-variants, each inclusive of the mcr-1 variants (Figure 3b, Table S3). Most mcr genes have been reported in studies from China (In Figure 3C), which is due to the large volumes of articles being published on mcr epidemiology from China.
It has thereafter been seen in China, that clones within mcr-positive E. coli (MPEC), which includes ST744, ST410, ST10, ST43, ST101 and ST206, have been identified in all four sources: food, environment, animals, and humans, seen in Table S4. These clones have, however, also disseminated globally, where ST744 has been identified in humans in eleven countries, in food in five countries, in animals in five countries and in the environment only in China. The MPEC ST10 strain is the most widely distributed within E. coli. It has been identified in animals in seventeen countries, humans in sixteen countries, the environment in five countries, and in food in five countries. MPEC clones within each country are usually found in both animals and humans, seen with ST744, ST69, ST117, ST131 and ST354 in Italy.
S. Typhimurium is also well distributed globally, with the S. Typhimurium ST34 strain identified in China, Colombia, Denmark, Germany, and the United Kingdom, in both animals and humans. In China, the S. Typhimurium ST34 strain has been identified in both humans and animals, and in food samples and human specimens in Germany. Similar results are seen in the monophasic variants of S. Typhimurium serovars (S.) 1,4,[5],12:i:-, and S. 4,[5],12:i:-, where ST34 was the only clone identified with mcr genes. The S. 1,4,[5],12:i:- ST34 strain has only been identified in Portugal in animals, food and humans, and the S. 4,[5],12:i:- has been identified in Belgium, Canada, Italy, Switzerland and the United States in animals and humans (Table S4).
The direct transmission of mcr-positive Enterobacteriaceae (MCRPE) isolates from animals and humans is discussed in this review (See Risk factors) and the data seen in Table S4 highlights this route of transmission.
Geographical and host distribution of clones and ARGs
Similar to mcr genes (Figure 3c), other antimicrobial resistance genes (ARGs) are predominantly located in China (Figure 4), followed by Germany, Denmark, and England. Notably, several important ARGs are co-hosted by MCRPE isolates (Figure 3A). This is expected as colistin is used as a last-line antibiotic for multi-drug resistant (MDR) isolates. A Table showing the distribution of mcr-positive isolates hosting other ARGs is shown in Table S5.

Global map showing the geographical distribution of antibiotic resistance genes found in all MCRPE.
Plasmid incompatibility groups associated with mcr genes
Mcr genes are usually associated with insertion sequences (ISs), which aid in the mobilisation of resistance genes from the chromosome to plasmids and vice versa 32. The dissemination of mcr genes is, however, mediated by plasmids, which allow for the horizontal transfer and spread of resistance genes across different bacterial species, genera, and families 33–35. Mcr genes have been identified with 41 different plasmid replicons, where eight of these are made up of IncHI2 or IncHI2A. Most mcr variants are associated with multiple incompatibility groups, with mcr-1 and its variants being associated with 37 plasmids; IncI2, IncX4 and IncHI2 were the most commonly reported (Figure 5). These incompatibility groups have also been shown to harbour other mcr variants: IncI2 has been associated with mcr-7, IncX4 with mcr-2, and IncHI2 with mcr-9, mcr-3, and its variants (Figure 5 and Table S6).

The distribution of plasmid incompatibility groups associated with each mcr gene.
Interestingly, the broad-spectrum IncHI2 plasmid was associated with the most mcr variants and is thus the main driver of mcr dissemination across different bacterial species. Further, mcr-2, mcr-6, and mcr-7 have only been identified in a single incompatibility group (Figure 5). This is because these mcr variants have only been identified in low numbers. A heat map (Table S4) shows the MGEs profile of the included isolates, which includes both plasmid groups and IS elements.
Discussion
Since the emergence of mcr-1 in 2015, it has spread from animals to humans through several sources and routes, mediated by chromosomal and plasmid borne mobile genetic elements. The genetic context, associated risk factors, protein structure, and enzymatic activity of each mcr variant are discussed herein.
Mcr-1 genomic content
The mcr-1 gene is part of a 2,600 bp cassette that is made up of a putative promoter gene responsible for the expression of the mcr-1 gene and the hypothetical protein later identified as pap2 36, 37. mcr-1 is speculated to have been derived from Moraxella species, which harbours the intrinsic chromosomal encoded mcr-like genes and the pap2 membrane-associated lipid phosphatase 37, 38. The pap2 gene is found in both mcr-1 and mcr-2 cassettes and shares 41% identity with Moraxella oloensis phosphatidic acid phosphatase 20, 39. Moraxella mcr-like genes with a significant degree of similarity to mcr-1 and mcr-2 in M. porci and M. osloensis were respectively identified in Genbank. Thus, these genes could be closely related 38. Poirel et al. (2017a) identified an mcr-like gene, mcr-2.2, from an M. pluranimalium strain with an 82% and 99% amino acid identity to mcr-1 and mcr-2, respectively.
An analysis of mcr-1 sequences deposited in GenBank in 2017 revealed an mcr-1.10 variant from Moraxella sp. MSGI3-CO3 with 97.61% identity to the plasmid-borne mcr-1 gene. This isolate was isolated from the faecal contents of healthy pigs in the United Kingdom in April, 2014 40. This data suggests that Moraxella species may have been the likely source of mcr-1 and mcr-237, 39. Other evidence that supports the speculation that mcr-1 evolved from the Moraxella species is the identification of an ISApl1 element in M. bovoculi and M. porci 39, 41. Li et al. (2018) suggest ISApl1 integrated into M. bovoculi and thereafter evolved with the mcr-like genes to the point seen today. This synteny of mcr-pap2 genes across Moraxella species further highlights this genus as a natural reservoir of mcr-like genes and a possible progenitor due to the high amino acid identity 38, 39, 42.
The mobilization of the mcr-pap2 unit was thereafter accomplished through IS elements, but Kieffer et al. (2017) identified a replicase gene associated with mcr-1 on IncX4 plasmids. The gene had a 99% identity to M. lacunata, thus showing this species as a possible reservoir of IncX4 plasmids and further speculating that the Moraxella family may encode genetic tools likely involved in the initial mobilisation of mcr genes 38.
Stoesser et al. (2016) and Sun et al. (2018) suggested that the initial mobilisation of mcr-1 genes into Enterobacteriaceae was IS1294-mediated, using a one-ended rolling circle transposition mechanism shown to be capable of mobilising adjacent sequences. The IS1294 may have mobilised the mcr-1 cassette into an ISApl1 composite transposon, creating an ISApl1-mcr-1-pap2-IS1294-pap2-ISApl1 cassette, which has been identified on E. coli chromosome 32, 43. It was found that the cassette was still flexible enough to jump from the chromosome to a plasmid 32 by generating a putative circular intermediate product.
There is increasing evidence, however, that the mcr-1 gene is mobilized primarily as a composite transposon, Tn6330, that is made up of two copies of ISApl1 that bracket cassettes 44, 45. ISApl1 is an IS that was first described in Acinetobacillus pleuropneumoniae and is part of the IS30 family 46. The IS elements of this family are flanked by 20-30-base pairs (bp) inverted repeats (left IR (IRL), right IR (IRR)), which are essential for transposition 47. The IR contains a 924 bp open-reading frame that encodes a 44.3 kDa transposase protein containing a DDE domain, which encodes three conserved amino acid residues viz., D228, D295, and E648 (DDE), as well as carboxylase residues that help coordinate metal ions for catalysis 44, 47. Analysis of each ISApl1 element flanking the mcr-1-pap2 unit found conserved dinucleotides between the ISApl1 inverted repeats and the mcr-1-pap2 unit, an AT dinucleotide on the IRR of the upstream element and CG on the IRL of the downstream element 44. An interesting observation in the mcr-1.10 variant identified in Moraxella sp. MSGI3-CO3 was the presence of these dinucleotides, AT upstream and CG downstream, flanking the mcr-1 structure 40. These dinucleotides were suggested to represent the ancestral target-site duplications (TSDs) formed during the initial mobilisation of mcr-1 during ISApl1 insertion 40. Therefore, Snesrud et al. (2018) suggested that the formation of the composite transposon, Tn6330, was through two independent insertion events of ISApl1 into the TA- rich region of the mcr-1-pap2 unit, generating the conserved interior 2bp TSDs, AT and CG 44. Subsequent transposition of the Tn6330 would therefore generate new TSDs at the new target site but would retain both internal conserved 2bp dinucleotides 40.
As stated above, the ISApl1 is part of the IS30 family. The family has been shown to mobilize through a copy (out) and paste mechanism, forming circular intermediates of a single IS during transposition. The family is further known to have a high affinity for certain target sites resembling their IR sequence 46–48. Multiple studies have investigated the mechanisms of ISApl1 in mobilising the mcr-1 gene and found that during each transposition event, the transposon was a circular intermediate covalently closed doubled stranded DNA, 5 699 bp in size and generated a 2 bp direct repeats at the insertion site which was an AT-rich region 37, 44, 47, 49, a similar mechanism seen in the IS30 family. ISApl1 is most likely an important factor responsible for the insertion and fixation of the mcr-1 gene into various classes of self-transmissible plasmids and host chromosomes 37, 41, 50. The formation of an intermediate structure consisting of ISApl1 and mcr-1 during mobilization indicates that the resistance genes have become highly mobilizable in both plasmids and the chromosomes 41. After the first identification of mcr-1 in pHNSHP45 11, an Incl2 plasmid, mcr-1 was thereafter detected in a wide range of conjugative plasmids, IncI2, IncHI2, IncX4, IncF, and IncP with the potential to mediate the dissemination of mcr-1 genes into other Gram-negative bacteria 50. Petrillo et al. (2016) suggest that the insertion of the complete transposon triggers the rapid mobilization of conjugative plasmids, encouraging their dissemination across the Enterobacteriaceae family 51. The copy out and paste in mechanisms allow for the transposition of the mcr-1 cassette, and the decay properties of ISApl1 further transfix the resistance gene into a plasmid or chromosome49, 52.
Snesrud et al. (2017) and Li et al. (2019) discovered that ISApl1 is highly active and that a single copy of ISApl1 can mobilize independently of mcr-1 across the host genome in AT regions with a slight central GC bias 49, 53. A sequencing analysis of four mcr-1 containing isolates performed by Snesrud et al. (2017) identified two to six copies of ISApl1 element throughout the isolates’ genome. The highly active nature of the ISApl1 elements thereafter triggers the deletion of the flanking ISApl1 copies to prevent further plasmid rearrangements 40, 49. Snesrud et al. (2016) analysis of the mcr-1 sequence environment showed that Tn6330 has a strong tendency to decay through deletion, removing parts of, or both copies of ISApl1, thus transfixing mcr-1 into a vector plasmid. This has led to the observation of many sequences lacking one or both of ISApl1 37, 44. Composite transposons in the IS30 family have been shown to contribute to replicon stabilization through transposition and illegitimate recombination 44, 47. The loss of ISApl1 elements results in the loss of transposability, stabilizing the mcr-1 cassette in plasmids, which facilitates the widespread dissemination of the colistin resistance gene in self-transmissible plasmids 44, 54, 55. As discussed, ISApl1 has a significant bias for insertion in AT-regions and generates TSD of two or three bases. The analysis of the mcr-1 cassette in the absence of ISApl1 elements has shown that the cassette is found in similar locations as per plasmid type and flanked by conserved trinucleotides (5’-ATA-3’) that are found immediately downstream of the ISApl1 IRR 44, 56. This is because the deletion event involves 1-4 flanking nucleotides that remain at the deletion junction 40.
An analysis of mcr-1 sequences deposited in the public database has shown four general structures of mcr-1 sequences: the complete composite transposon with both copies of ISApl1 elements; structures with a single copy of ISApl1 located downstream of mcr-1; structures that lost both copies and a rare fourth structure with a single copy of ISApl1 located upstream 40, 44. In structures with a single copy of ISApl1 found upstream and the IRR sequence of the downstream of deleted ISApl1, Snesrud et al. (2016) suggest that the transposase encoded by the upstream ISApl1 can recognize the downstream IRR and thereafter still be able to mobilize the bracketed region without a complete composite transposon 44. The partially or complete removal of ISApl1 was through an illegitimate recombination that generated mismatches and deletions. Sun et al. (2018) found that the 3’ end of the mcr cassette unit was flexible in all IncX4 plasmids and the sequence could match with the perfect IRR of ISApl1, though all IncX4 currently lack ISApl1 elements. The evidence from this study suggests that the TSD generated, and the six mismatches acquired through illegitimate recombination could be identified as a “relic” to track an insertion event that resulted in the subsequent loss of ISApl1 32. The loss of ISApl1 elements in IncX4 was conducive to maintaining the mcr-1 cassette on the plasmid 32, increasing its stability, and thus allowing for the dissemination of resistance genes via the plasmids 44, 54, 55. The identification of these IRR in IncX4 plasmids allows for the conclusion that ISApl1 was associated with the transposition of the mcr-1 cassette into IncX4 plasmids 57.
Fitness cost of mcr-1 genes on bacterial host
The acquisition of mcr-1 bearing plasmids has been shown to have a beneficial effect on the host, improving bacterial survival in the presence of colistin treatment 58. The acquisition and expression of the mcr-1 gene results in the incorporation of MCR-1 into the bacterial membrane and the phosphoethanolamine (PEtN) modification of the lipopolysaccharides (LPS) 59. However, multiple studies have shown that the expression of mcr-1 imposes a fitness cost by placing an energy burden on the host 60, impairing cell growth, and diminishing bacterial fitness 59. Andersson et al. (2006) 61 explained that a significant fitness cost is seen when the susceptible strain outcompetes the resistant strain in an antibiotic free environment. The imposed fitness cost of a conjugative plasmid harbouring a resistance gene has been shown to be because of various factors such as the resistance mechanisms, the bacterial species, and the antibiotic 58.
Evaluating the cost of colistin resistance on resistant strains, it is shown that certain chromosomal mutations within genes such as mgrB have no significant fitness cost on K. pneumoniae 62 whilst genes such as pmrB, do 63. Giordano et al. (2019) 63 found that the expression of the mcr-1 gene imposes less of a burden on K. pneumoniae than the mutated pmrB gene. The chromosomal mutations within pmrAB, crab, phoPQ, and mgrB genes have been shown to mediate colistin resistance through the phosphorylation of lipid A 64–66. Tietgen et al. (2018) discovered that the fitness costs imposed by mcr-1 plasmid carriage, such as growth rates and cytotoxicity, could be species-specific 67. Particularly, the acquisition of an mcr-1-harbouring plasmid in K. pneumoniae has been shown to impose a significant fitness cost on the host 58, but multiple studies have shown that the acquisition of an IncI2 plasmid, in various sizes, carrying mcr-1 had no significant cost on the host 68, 69. This may be plasmid and/or species specific or may be due to acquired compensatory mutations within the IncI2 plasmid. However, the overexpression of mcr-1 was seen to result in profound changes in the architecture of the outer membrane, resulting in the loss of membrane structural integrity and causing leakage of cellular cytoplasm, resulting in cell death 59, 70.
Yang et al. (2017a) 59, 70 found that the overexpression of mcr-1 imposes a significant cost by decreasing the growth rate, causing significant membrane degradation and moderate fitness loss. They came to this conclusion through evaluating the effects of mcr-1 expression on the relative fitness of E. coli TOP10 and found that with increasing levels of mcr-1 expression resulted in a significant fitness burden on the host 59, 70. An analysis of the cellular morphology in the mcr-1 overexpressed strains using a transmission electron microscopy showed cell architecture alterations and a complete loss of cellular morphology. The overexpression of mcr-1 altered the structural integrity of the outer membrane and further impaired the cell membrane 59. They finally found that the embedding of MCR-1 into the outer membrane and the PEtN modifications of Lipid A were the leading factors contributing to fitness cost and membrane degradation. Therefore, the expression of mcr-1 in host strains is tightly controlled to regulate the mcr-1 fitness cost 70.
Mcr-1 expression is therefore quite toxic for the bacterial host, and the acquisition of a mcr-1- bearing plasmid thereafter results in compensatory adaptation that allows for the maintenance of high-cost conjugative plasmids 19, 71, 72. Dahlberg et al. (2003) 72 found that the cost of plasmid carriage is reduced over long-term culture because of compensatory mutations. The host chromosome or plasmid evolves compensatory mutations that, in the case of plasmids, enhance the fitness of the host, and in the case of chromosomal mutations, aid the host in evolving towards new growth conditions by decreasing the plasmid carriage cost 72. This has been seen in multiple studies where mcr-1 bearing plasmids initially imposed a biological cost on the transformant. However, overtime, the cost of plasmid carriage in long-term cultures was largely compensated for and plasmids were stably maintained through passages 59, 63, 67, 71, 73.
Ma et al. (2018a) 71 investigated the potential mechanisms involved in compensatory adaptation through comparative genomics and identified single nucleotide polymorphisms in several genes. Amongst the genes identified in this study are dnaK, which encodes a molecular chaperon involved in chromosomal DNA replication, and cpoB, which encodes an RHS repeat protein involved in maintaining the cell envelope integrity during cellular division 71. These two genes are located on the chromosome and have non-synonymous single nucleotide polymorphisms (SNPs). The role of these genes in reducing plasmid carriage cost is unknown and may represent novel mechanisms. The study further shows that bacteria may use different strategies to reduce the fitness cost of plasmid carriage under different environmental conditions 71
In the absence of antibiotics (colistin) selective pressure, mcr-1 bearing plasmids have been shown to be less maintained and that the complete elimination of the mcr-1 gene within a population is possible 58. Nang et al. (2018) 58 performed a plasmid stability assay and found that in the absence of colistin, there was a gradual loss of the mcr-1 plasmid, there was a decrease in its maintenance within the population. This may be due to the instability of mcr-1 harbouring plasmids 58. Arcilla et al. (2016) have shown the complete elimination of mcr-1 bearing bacteria in travellers returning to their home country after a month 74.
MCR-1 structure
MCR-1 is a 541 amino acid, integral membrane protein made up of two domains: a C-terminal periplasmic catalytic domain and an N-terminal 5’-helix transmembrane domain 75, 76. The transmembrane (TM) domain is made up of 5-membrane spanning α-helixes with an overall positive charge because of positively charged residues, which interact with the negatively charged phospholipid head groups of the membrane bilayer. The TM domain anchors the protein into the inner membrane and is connected to the catalytic domain through a bridging helix 75. The catalytic domain is made up of both positively and negatively charged residues, where the negatively charged residues create the binding pockets and allow them to be buried within the domain 75.
The overall shape of MCR-1 is a hemispherical shape composed of several β-α-β-α motifs made up of β-strands sandwiched between α-helical structures 12, 77. The catalytic domain has an alkaline phosphatase family α/β/α fold, also with a hemispheric shape equipped with a zinc binding pocket containing a conserved phosphothreonine-285 residue (Figure 6)12, 77. The pocket is common to PEtN transferases and alkaline phosphatase; each enzyme, however, differs in the orientation and number of zinc molecules 77. The pocket is proposed to be critical for the nucleophilic attack of the phosphate of the donor PE substrate by stabilizing the alkoxide of the Thr285 77. The Thr285 residue, is the catalytic nucleophile that acts as both a nucleophile and a PEtN acceptor during the catalytic mechanisms. The residue is critical to MCR-1’s function as mutations to this residue significantly decrease MCR-1’s activity 77, 78. The catalytic domain of MCR-1 is further made up of six cysteine residues that form three disulphide bonds, Cys281/Cys291, Cys356/Cys364 and Cys414/Cys422 77 that stabilize and anchor the β-α-β-α motifs 12. These disulphide bonds are conserved, and equivalents are present in both LptA and EptC transferases 77, although LptA has four more cysteine residues.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Structure of the catalytic domain of the MCR-1 protein and the five important residues required for its catalytic activity. A) The MCR-1’s hemispherical shape composed of the β-α-β fold made up of helices (cyan), strands (purple) and loops (salmon). Image was obtained from Stojanoski et al. (2016)209. B) A ball and stick model showing the conserved active site residues of MCR-1 consisting of Asp (D)465, Glu (E)246, His (H)466, His(H)395 and Thr (T)285, coordinated to the zinc ions (Zn1 and Zn2) and water molecules (W1 and W2). Image was obtained from Ma et al. (2016) 210.
The zinc binding pocket is in the active site of MCR-1 and is made up of conserved residues viz., Asp465, Glu246, His466, His395, and Thr285, and is made up of zinc molecules (Figure 6b)77, 79. The conserved residues are critical for the substrate binding of MCR-1 79 and mutations in these amino acids abolish the MCR-1 activity, lowering the colistin MIC value down to control levels 13. The five conserved residues are well conserved in MCR-1, LptA, EptC and other alkaline phosphatase family members, although the nucleophile residue may differ between enzymes 78. MCR-1 has three zinc molecules clustered around the active site: Zn1 is buried within the active site and co-ordinated by the phosphate oxygen of Thr285; it is conserved in LptA and EptC 77. Zn2 is co-ordinated by His395, bound to a phosphate oxygen of Thr285 and three water molecules, forming a trigonal bipyramidal configuration seen in Figure 6b 77. Hu et al. (2016a) found that Zn2 is not critical for maintaining MCR-1 activity.
This may be because it is located on the surface of the active site. A structural comparison between LptA, EptC and MCR-1 found that the Zn2 site is less conserved. Hence, Hinchliffe et al. (2017) 78 concluded that an intact Zn2 site was not a prerequisite for MCR-1’s catalytic activity. However, His395 residue is important for the structure and activity of MCR-1. The Zn3 molecule, unlike the other zinc molecules, is not co-ordinated by the conserved residues but is tetrahedrally co-ordinated by four water molecules. The seven water molecules found in the active site, co-ordinating Zn2 and Zn3, are embedded in the protein through hydrogen bonding 77. The catalytic domain is thus a zinc-rich area (Fig. 6b), suggesting that MCR-1 may be able to attract zinc ions at different levels into the domain 13. The role of zinc in mediating lipid A modification, the catalytic mechanism, was evaluated by measuring the colistin MIC in the presence and absence of EDTA. Hinchliffe et al. (2017) 78 found clear reductions in colistin MIC values after EDTA treatment and concluded that the presence of zinc in the active site was important for MCR-1 function.
MCR-1 mediates the transfer of PEtN from the primary phosphatidylethanolamine (PE) to lipid A moiety at the 4’-phosphate group 13, 79. Wei et al. (2018) 75 and Liu et al. (2018) 76 investigated the molecular mechanism involved in this process. Wei et al. (2018) identified that the active site within the catalytic domain is made up of two binding pockets: the PEtN binding pocket and the Lipid A binding pocket. These binding pockets are in close proximity and are integrated within each other 75. Wei et al. (2018) 75observed that ethanolamine was a good analogue of PEtN and D-glucose for lipid A. Ethanolamine (ETA) is a good analogue as it mimics the covalent PEtN-enzyme intermediate. Lipid A is made up of two glucosamine units that are attached to acyl chains. Thus, monosaccharides and disaccharides can mimic a Lipid A molecule as they are both hexacyclic compounds 75, 76.
The PEtN binding pocket that accommodates ETA is made up of Glu246, Thr285, Asn329, Lys333, His395, Asp465, His466 and His478 residues 75. This pocket is responsible for the binding of phosphatidyl-ethanolamine (PE), which is stabilized through hydrogen bonding with Asn329, a water molecule, and phosphorylated Thr285 75. Analysis of the entry of ETA into the pocket showed that the pocket undergoes conformational changes to adjust to the substrate. These included a 50° rotation of the His395, accomplished by breaking a hydrogen bond with a water molecule (wat7) and creating a new hydrogen bond with a new water molecule (wat8) 75. This releases the wat7 molecule from the PEtN binding pocket and thus, collectively, the conformational changes create room to accommodate the substrate, ETA. The entry of ETA further deprotonates the phosphorylated Thr285 residue, forming a PEtN molecule and a nucleophilic state at Thr285 75. The Zn1 molecule subsequently facilitates the stabilization of the nucleophilic state of Thr285 that allows that residue to thereafter attack the PEtN, creating a PEtN enzyme intermediate. This constitutes the first step of the MCR-1-mediated catalytic process 75.
Wei et al. (2018) 75 and Liu et al. (2018) 76 investigated the mechanisms behind PEtN-transfer reaction with a lipid A analogue, D-glucose, identified within the Lipid A binding pocket. This pocket is made up of the following residues: Thr283, Ser284, Tyr287, Pro481 and Asn482, that is located near the Thr285 residue (Figure 7). The analysis of D-glucose found that D-glucose was held in the pocket by Thr285, Ser284 and Asn482 and was flanked by Tyr281 and Pro481 forming a sandwich structure of a π-π conjugative interaction 75, 76.The importance of the residues in both PEtN and Lipid A binding pockets in MCR-1 activity were evaluated through mutation construction. Mutations within Thr285, Asn329, Lys333, Glu246, His398, Asp465, His466 and His478 of the PEtN binding pocket and Tyr281 and Pro481 of the Lipid A binding pocket, resulted in a decrease in colistin MIC values 75. However, these mutations did not disrupt the protein expression and membrane localization of the MCR-1 protein. The mutations of Tyr281 and Pro481 in the lipid A binding pocket highlights the importance of this pocket in the MCR-1 activity. Wei et al. (2018) 75 suggests that it could bind to the glucosamine group of Lipid A and allows for the transfer of the reactive phosphate group from PEtN enzyme intermediate to lipid A. Thus, the second step of the catalytic process therefore involves the transfer of PEtN to Lipid A that is situated in a nearby pocket 75.
The MCR-1 catalytic domain is therefore made up of a PEtN binding pocket and a Lipid A binding pocket that mediate the transfer of PEtN from the PEtN binding pocket to lipid A moiety at the 4’-phosphate group 13, 79. This activity is validated through MALDI-TOF mass spectrometry (MS), provides in-vivo evidence of the MCR-1 activity 79, 80. This is accomplished by evaluating the LPS lipid A MS profile in the presence and absence of mcr-1 expression as adding PEtN increases Lipid A mass units (+123) 79, 81, 82. In the absence of mcr-1 expression, there is a single Lipid A peak (m/z = 1797.4) and in the presence of mcr-1 expression, there are two unique peaks. The first peak is the wildtype bis-phosphorylated, hexa-acylated lipid A (m/z = 1797.4) and the second peak is the PPEtN-4’ (m/z = 1920.5) 81.
MCR-1 has thus proven to have PEtN transferase activity and has a sequence and structural similarity to EptC and LptA PEtN transferases in Campylobacter jejuni 14 and Neisseria meningitidis 83, respectively 77–79. The active sites of the three PEtN transferases are highly conserved, i.e., the phosphorylated catalytic nucleophile Thr285, disulphide bonds, zinc binding pockets and the conserved active site residues 78. Therefore, these three sites are important for MCR-1 function; these conserved mechanistic activities and binding pockets will allow for the use of MCR-1 inhibitors to also inhibit chromosomally encoded PEtN transferases, restoring colistin susceptibility in acquired and intrinsically resistant bacteria 77.
Mcr-2 variant
Mcr-2 was first identified in ten E. coli isolates during a passive surveillance screening of diarrhoea in calves and piglets, and like mcr-1, mcr-2 was located on a conjugative plasmid (IncX4) and was able to confer clinical colistin resistance (4-8 mg/L) 20. The gene was identified due to the presence of a putative membrane protein identified as a PEtN transferase in mcr-1 negative isolates 20. This variant has a 76.75% and 80.65 % nucleotide sequence and amino acid identity, respectively, to mcr-1 20. mcr-2 encodes a 538 amino acid polypeptide consisting of two domains: a C-terminal periplasmic catalytic domain and an N-terminal 5’-helix transmembrane domain 20, 33. Sun et al. (2017b) found that the acquisition of the transmembrane domain allows for the correct localization of PEtN transferases within the periplasm and that the deletion of this domain impairs MCR-2’s ability to confer colistin resistance.
A genetic analysis of the IncX4 plasmid found that mcr-2 was located within two IS1595-like ISs and a 279 bp open reading frame (ORF), located downstream the mcr-2 gene 20. The ORF encodes a PAP2 membrane-associated lipid phosphatase which is related to the PAP2 protein encoded by the ORF of mcr-1 20, 84. The ORF of mcr-2 has a 41% identity to the phosphatidic acid phosphatase encoded by M. osloensis. When Xavier et al. (2016) removed the mcr-2-pap2 from the IS1595 backbone, a BLASTn search produced a single hit to M. bovoculi strain with a 75% identity across a 100% query coverage. To support the hypothesis that the mcr-2 cassette originated from Moraxella sp., a phylogenetic analysis of the MCR-2 protein was performed and found that the protein was a distinct protein from MCR-1 and might have evolved from M. catarrhalis 20. Sun et al. (2017b) suggests that the emergence of the PEtN transferases, MCR-1, MCR-2 and Neisseria LptA was a parallel evolutionary event for functional acquisition of colistin resistance during some environmental selection pressure, i.e., the intensive use of colistin in animal feed 33.
IS1595 is composed of a transposase gene flanked by two inverted repeats (18 bp each). Mcr-2 is thus present within a complete transposon which allows for horizontal gene transfer 33. Sun et al. (2017b) found that the IS1595 composite transposon was able to form circularized intermediates, which were identified during an inverse PCR assay. Hence, the IS1595 elements are involved in the mobilisation of the mcr-2 cassette via homologous recombination events, which like mcr-1, allows for the dissemination of the resistance gene across diversified bacterial hosts 33.
After the first identification of mcr-2 in Belgium on an IncX4 plasmid, an effective vehicle for dissemination of resistance and virulence genes with a high transfer frequency 20, 33, it was thereafter identified in Salmonella sp. on IncX4 plasmids in Belgium in 2018 from retail meat collected in 2012 85. It has subsequently been found in pig and poultry samples in 19 provinces in China, where the prevalence of mcr-2 in the study was 56.3% in poultry 86. Mcr-2 was then found in clinical settings in Iran, in stool samples collected between 2011-2016 87. In Egypt, mcr-2 was widespread and identified in humans, wild birds, and the environment (water sources) 88. Compared to mcr-1, mcr-2 has not spread across the world and imposed a big threat to public health though found on a low fitness burden plasmid. The gene has however spread well within the Enterobacteriaceae family i.e., E. coli, K. oxytoca, K. pneumoniae and Salmonella sp.
The mcr-2.2 variant: mcr-6
AbuOun et al. (2017)42 identified an mcr-2.2 variant in Moraxella pluranimalium strain isolated from a pig in Great Britain. The MCR-2.2 protein was found to have an 87.9% amino acid identity to MCR-2, encoding 65 amino acid substitutions 42. Partridge et al. (2018) 89 suggested that since mcr-2.2 encodes a protein that is 87.9% identical to the original MCR-2, it should be labelled as mcr-6. The mcr-6 gene like mcr-1 and mcr-2 was associated with the PAP2 gene located downstream of mcr-6 42. The mcr-6 has only been identified in the M. pluranimalium MSG47-C17 strain isolated in 2012 and has not disseminated into the Enterobacteriaceae family. AbuOun et al. (2017) 42 however highlights that Moraxella species could have been the source of mcr-pap2 genes.
Mcr-3 variant
The first identification of mcr-3 was in E. coli isolated from pigs in China 90. The identified 1626 bp putative PEtN transferase has a 45.0% and 47.0% nucleotide sequence identity to mcr-1 and mcr-2 and an amino acid sequence identity of 32.5% and 31.7%, respectively 90. Using RaptorX, Yin et al. (2017) 90 predicted that MCR-3 protein, like MCR-1, and MCR-2, is made up of two Domains, domain 1 containing a five-transmembrane helix and domain 2 containing the periplasmic domain made up of the putative catalytic centre 90. Despite the difference between mcr-3 and the other mcr genes, Kieffer et al. (2018) 91 found that an mcr-3.12 variant had the same PEtN transferase activity and the expression of mcr-3 resulted in an 8-fold increase in MIC value of colistin 92. An LPS analysis showed that both MCR-1 and MCR-3 produced an identical additional peak at m /z 1921 in mcr-1 and mcr-3 positive E. coli. Thus, the differences in nucleotide or amino acid sequences between the two mcr genes had no impact on the PEtN activity91, 93 and proving that MCR-3 confers colistin resistance in the same manner as MCR-1 and MCR-2 91.
To further elaborate this observation, the expression of mcr-3 has been shown to significantly impair the cell wall integrity and decrease the electron density of mcr-3 producing E. coli 94, a characteristic that has been previously described in mcr-1 producing isolates 67, 70, 94. Yang et al. (2020) 94 further evaluated the fitness cost imposed by mcr-3 expression and compared it to that of mcr-1 and found that the expression of both mcr genes had an impact on the bacterial fitness; however, the mcr-3.1 and mcr-3.5 imposed lesser fitness costs.
Mcr-3 genetic context is made up of a diacylglycerol kinase (dgkA) gene located downstream the mcr-3 gene and is thereafter flanked by a truncated (Δ) ISKpn40 upstream and an intact ISKpn40 downstream 52. In some cases, a ΔTnAs2 element is located upstream of the ΔISKpn40 element 52, 90, 95, and the genetic context (ΔTnAs2-ΔISKpn40-mcr-3-dgkAISKpn40) is flanked by an IS26 element upstream and an intact IS15DI element downstream96,34, 52. These elements, ISKpn40, TnAs2, IS26 and IS15DI were identified and hypothesised to play a role in the transposition of the mcr-3 cassette among different species or bacterial genera 35, 52, 97. Sia et al. (2020)52 identified two circular intermediates of 3535 bp and 5990 bp made up of mcr-3-dgkA-ISKpn40 and ISΔ26-TnAs2-ΔISKpn40-mcr-3-dgkAISKpn40, respectively, during conjugation experiments. Wang et al. (2018) 35 evaluated the transferability of the 5990 bp circular intermediate and concluded that the ΔIS26 and intact IS15DI mobilises the mcr-3.1 via homologous recombination through the formation of circular intermediates. The IS-mediated transposition enables the mobilisation of the mcr-3 resistance gene between the chromosome and plasmids, which further contributes to the dissemination of resistance genes 34, 35. Similar conclusion was reached with the 3535 bp circular intermediates in a study performed by Xiang et al. (2018), 97 which showed that mobilisation of the mcr-3 core segment (mcr-3-dgkA) through ISKpn40 elements facilitated its spread into various plasmids.
The mcr-3.12 variant has a 99% nucleotide sequence identity with an Aeromonas veronii sequence 91 and significant (77%) amino acid sequence identity to three PEtN transferases found within the genera 90, 91, 98. It is suggested that mcr-3 genes originated from the Aeromonas genus, or the mcr-3 gene is widely disseminated as an acquired resistance trait and thus also found in Aeromonads 91. It is however thought that mcr-3 was initially transposed from the Aeromonas genera to the Enterobacteriaceae family through ISKpn40- mediated transposition. The ISKpn40 elements encode two ORF and the second ORF has a 99% identity to both a transposase from Aeromonas sp. and an integrase from Aeromonas caviae 99. This IS element is found flanking the mcr-3 segment and was also shown to mobilise through homologous recombination 52, 97. The last evidence of mcr-3 originating from Aeromonas includes the presence of TnAs2 located upstream of the mcr-3 gene 90. The transposon has only been identified in A. salmonicida and because its sequence in pWJ1 (original mcr-3 plasmid) is a partial sequence, it is unlikely to have mobilised the mcr-3 gene 90, 95. This evidence supports the possibility of the Aeromonas genus being a possible progenitor of mcr-3 genes 90.
Mcr-3 and its variants are disseminated within Enterobacteriaceae, having been identified in E. coli, Salmonella sp. and K. pneumoniae in healthcare centres, aquaculture, wastewater, and in animals 18, 34, 90, 92, 95, 98, 100. The resistance genes have been identified in both transmissible and non-transmissible plasmids such as IncA/C2, IncHI2, IncHI2A, IncFII/FIB, IncY, IncR, IncF, and IncP and in many variant mutations. The first identified mcr-3 gene was identified on an IncHI2 plasmid replicon (pWJ1) in E. coli isolated from pigs in China 90. Thereafter, multiple mcr-3 variants have been identified with one or more amino acid substitutions. Yang et al. (2020) evaluated the impact of mcr-3 expression on bacterial fitness using two mcr-3 variants, mcr-3.1 and mcr-3.5 (T488I, M23V, A456E) 94. The study showed that although the expression of both variants imposed a fitness cost and impaired the cell wall integrity, the expression of mcr-3.5 was less costly 94. Yang et al. (2020) 94 showed that the amino acid substitutions A457V and T448I, seen in mcr-3.5, had strong compensatory effects when introduced in mcr-3.1. The introduction of these substitutions, individually, resulted in an increased fitness of up to 45%. However, double substitutions demonstrated a negative epistasis.
This study highlights an interesting concept about all mcr genes i.e., the mcr variants may encode compensatory mutations that mitigate the expression of mcr genes and thus, is an evolutionary mechanism for the worldwide dissemination of mcr genes 94. Though variants such as mcr-3.10 has seven substitutions (V122G, R297L, E337K, H341Y, D358E, Q468K and I313V) 92 in both the putative transmembrane region and catalytic domains, these substitutions do not affect MCR-3.10’s ability to confer colistin resistance. Wang et al. (2018d) showed that similar to the original mcr-3, the expression of mcr-3.10, results in an 8-fold increase in colistin MICs. Similar results were seen with mcr-3.5 95.
Mcr-4 variant
Mcr-4 was first identified in S. Typhimurium isolated from the caecal content of a pig at slaughter in Italy in 2013 101. The S. Typhimurium was colistin-resistant and negative for known mcr genes. MCR-4 respectively has an amino acid sequence identity of 34%, 35% and 49% to MCR-1, MCR-2, and MCR-3, and like other PEtN transferases, can mediate colistin resistance through Lipid A modification 101. MCR-4, however, has an 82%-99% amino acid sequence identity to one large PEtN transferase found in Shewanella sp., and the mcr-4.3 variant has a 100% nucleotide sequence identity to a chromosomal PEtN transferase encoded by S. frigidimarina 98, 101, 102. Mcr-3 and mcr-4 are suggested to have originated from aquatic environments, as Aeromonas and Shewanella sp. are aquaculture fish pathogens that are intrinsically resistant to colistin 98. The dissemination of these mcr genes to the Enterobacteriaceae family may have been carried through plasmids from the host genome. The mechanism underlining this mobilisation is yet to be discovered 98.
Mcr-4 was initially identified on an 8749 bp ColE10 plasmid, which encoded a RepB replicase, mobA/L mobilisation proteins, a RelE toxin and excl1 gene 101, 103. In other cases, the ColE plasmid was found to encode a RepA replicase instead of RepB. However, both plasmids showed a 99% nucleotide sequence identity to plasmids from Pantoea species, with a 65% coverage 101–103. The mcr-4 gene was flanked by IS5 elements on some plasmids and on other plasmids, it was flanked by an ΔIS10 element upstream and an IS1294 element downstream 103. Genetic analysis found that the ColE plasmid encodes a conserved 59-bp region upstream the mcr-4 gene, and this region is predicted to encode the -35 (TTATTT) and -10 (AGCTAGTAT) promoter regions.102 This allows for ColE plasmids to replicate independently in different bacterial species and genera 101. ColE plasmids are broad-range non-conjugative plasmids whose mobilisation, however, require a helper plasmid; nevertheless, the mobilisation of the mcr-4 gene is hypothesized to be achieved through a transposition event mediated by the IS5 element 101.
A genomic analysis of the initial mcr-4-producing S. Typhimurium genome found the ColE10 replicon located within the host chromosome integrated within the Type 1 methylation gene. Carattoll et al. (2017) suggest a transposition event mediated the chromosomal integration of the ColE10 plasmid 101. Mcr-4 has been isolated from both humans and animals in Spain, Belgium, Italy, and China in Enterobacteriaceae101, 104–106. An inactive mcr-4.3 variant identified by Teo et al. 105 (2018) was found to encode two missense mutations, V179G and V236F, that inactivated the PEtN transferase activity 102, 105. A comparative alignment of the MCR-4.3 to MCR-1 and MCR-2 found that the active residues were conserved in MCR-4.3 105. However, an MS spectrum of lipid A in mcr-4.3-expressing isolates showed mcr-4.3 failed to modify lipid A 102. The transformation of E. coli with an mcr-4.3 expressing vector resulted in no difference in colistin MIC values, suggesting that mcr-4.3 does not confer colistin resistance 102, 105. Thus, some compensatory mutations in mcr may aid in fitness cost while others such as those seen in mcr-4.3 significantly altered the PEtN transferase activity of mcr-4.3 102.
Mcr-5 variant
Mcr-5 was first identified in a d-tartrate-fermenting S. Paratyphi B isolated from food-producing animals across Germany between 2011 and 2013. Following this discovery of mcr-5, further screening resulted in the identification of fourteen additional mcr-5-producing isolates. These were isolated between 2011 to 2016 in the same study 107. Mcr-5 encodes a 547 amino acid PEtN transferase, which has a protein identity of 33% to 35% to other known MCR proteins i.e., MCR-1 to MCR-4. Although MCR-5 has a low identity to other MCR proteins, all five MCR proteins, i.e., MCR1, MCR-2, MCR-3, MCR-4, and MCR-5, each encode the five conserved residue viz., E248, T286, H389, D458, and H359, that are located within the catalytic centre of the periplasmic domain 107. These conserved residues have been found to be essential for substrate binding and MCR activity 79. The capability of mcr-5 to confer colistin resistance was shown using the original mcr-5-harbouring plasmid and a plasmid vector encoding the mcr-5 operon in a transformation experiment using both an E. coli and a wild-type S. Paratyphi strains. In both experiments, the transformants achieved a colistin MIC value of 8 mg/L 107.
The MCR-5 protein has a 100% amino acid sequence identity to a PEtN transferase identified in Pigmentiphaga genus and Cupriavidus gilardii. The first 404 amino acids of the MCR-5 protein also have a high identity to a hypothetical protein from Pseudomonas aeruginosa 107. Genetic analysis of mcr-5 found that the resistance gene is located within an operon encoding a Chromate B (ChrB) protein domain responsible for regulating the expression of ChrA for chromate resistance and two ORF that encode a major facilitator superfamily (MFS) type transporter 107, 108. The complete operon is located within a 7337bp Tn-3 type transposon named Tn6452 52, 107, that is flanked by inverted repeats (IRs). A genetic analysis of the transposon identified tnpA encoding a Tn3 transposon, tnpR encoding a Tn3 resolvase and a bla gene encoding a β-lactamase 109. Tn6452 encodes mechanisms for transposition of the mcr-5 cassette, TnpA, and TnpR 109, which allows the mcr-5 cassette to be transferred between plasmids and chromosomes 110.
Mcr-5 has also been identified on Tn3-like structures that encode the ChrB gene but lacked the transposase gene and encoded an imperfect Tn3-like inverted repeats left (IRL) 110–112. This structure resembles features of a miniature inverted repeat transposable element, which possess a left or right IRs but lacks a transposase gene 111. Kieffer et al. (2019a) 111 suggests the acquisition of the mcr-5 cassette was through a transposition event and although the Tn3- like structure lacks a transposase gene, it can still mobilise the cassette through a nonautonomous transposition mechanism. The mcr-5 cassette has been identified on the host chromosome 107, a multicopy ColE-type plasmid 107, 110, 112, 113 and on an IncX plasmid 110. The presence of mcr-5 cassette on a multicopy plasmid (ColE-type) thus means multiple copies of mcr-5 transposon will exist within the cell. Borowiak et al. (2017) 107 found that there is a higher degree of colistin resistance (8 mg/L) in isolates harbouring ColE-type plasmids encoding mcr-5 than in isolates with a single copy of the mcr-5 integrated within the chromosome (4 mg/L). The ColE type plasmids are, however, non-conjugative and thus the presence of the mcr-5 cassette on an IncX plasmid allows for horizontal transfer and dissemination of resistance gene between different bacterial genera and species 109, 111.
Mcr-7 variant
Mcr-7.1 was first identified in chicken in China 114. It has thereafter been identified in environmental samples 115, 116 and faecal samples in Brazil 117. The putative PEtN transferase has a 78% nucleotide sequence identity to part of the mcr-3 gene 114. Genetic analysis of mcr-7.1 identified a 381 bp ORF encoding the diacylglycerol kinase, dgkA, downstream of mcr-7.1. The dgkA has an 82% nucleotide sequence identity to Aeromonas 114. Yang et al. (2018) 114 suggests, like mcr-3, mcr-7.1 originates from Aeromanas. This was derived based on the close genetic distance between MCR-3 and MCR-7.1 found during a phylogenetic analysis and the presence of dgkA gene downstream both mcr-3 and mcr-7.1. The ORF sequence found upstream the mcr-7.1 gene has an 81% nucleotide sequence identity to a putative phosphodiesterase gene found in Aeromonas 114. This evidence suggests that the Aeromonas genus is a possible progenitor of mcr-7.1.
The mcr-7.1 encodes a 539 amino acid PEtN transferase that confers colistin resistance during conjugative experiments 114. The protein is made up of two domains, which resembles a similar structure of MCR-3. Mcr-7.1 was identified on a self-transmissible IncI2 type plasmid with no ISs in its environment; hence, the dissemination of mcr-7.1 may be achieved through plasmid mobilisation 114.
Mcr-8 variant
Mcr-8 was initially identified in K. pneumoniae from a swine faecal sample in China 118. Conjugation assays found that it is a functional PEtN transferase and its expression resulted in a four- and eight-fold increase in colistin MIC.118–120 MCR-8 mediated PEtN modification, which was identified using a MALDI-TOF-based technique that discriminates between colistin resistance mechanisms, i.e mcr-expression or chromosomal-mutations 120. BLASTn analysis showed that mcr-8 has a 50.23% nucleotide and 39.96% amino acid sequence identity to part of mcr-3 but has 30% to 40% amino acid sequence identity to other MCR proteins. Characterisation of MCR-8 using RaptorX predicted that MCR-8, like other MCR proteins, is made up of two domains: domain one consisting of a five transmembrane α-helix, and domain 2 consisting of the putative catalytic centre 118. Wang et al. (2018c) 118 performed sequence alignment of all eight MCR proteins and found that the active site residues (E246, T285, K333, H395, D465, H466, E468, H478) necessary for enzyme activity of MCR proteins and the six-cysteine residues required for the three disulphide bonds found in the catalytic domain were highly conserved across all 8 MCR proteins 118, 119.
Analysis of mcr-8’s genetic environment found that it is usually flanked by two complete IS903 elements, with both 50 bp IRL and IRR located upstream, and downstream, respectively 119, 121. It is also associated with ISEcl1 and ISKpn26, located upstream and downstream, respectively 122. Yang et al. (2019b) 122 found that the insertion of ISEcl7 was an independent event and may not be involved in mobilising mcr-8. The formation of circular intermediates via IS903B elements to mobilise mcr-8 remains unknown; however, the association of mcr-8 to ISs facilitates both the transmission of mcr-8 and its close association with other resistance genes 121; specifically, it has been found associated with aminoglycoside resistance genes 121, β-lactamase genes 119 and tmexCD1-toprJ1 genes that encode a novel plasmid-mediated efflux pump that confers resistance to tigecycline 123.
Mcr-8 is commonly found in K. pneumoniae 118, 120, 121, 123–126, Wang et al. (2018c)118 performed a phylogenetic analysis of the mcr-positive K. pneumoniae isolates identified in their study and found that the mcr-positive K. pneumoniae were genetically diverse. K. pneumoniae express colistin resistance usually through chromosomal mutations 10. Wu et al. (2020) 121 suggest the expression of mcr-8 may have synergistic effects with the chromosomal mutations within the two-component system (TCSs), resulting in heterogenous colistin resistance mechanisms 121. Mcr-8 has also been identified in Klebsiella quasipneumoniae 122 and R. ornithinolytica as well as on a diverse range of Inc groups: IncFII, IncFIA, IncFIB, IncQ, IncR, IncFIIK, IncFIA and IncA/C; these will expectedly accelerate the dissemination of mcr-8 and other ARGs within and outside Enterobacteriaceae 119, 121.
Mcr-9 variant
Mcr-9 gene was identified by Carroll et al. (2019) 127 in a colistin susceptible S. Typhimurium isolated from a human patient in Washington State in 2010. A BLAST analysis identified the gene in 355 genomes, where 65 were encoded on a plasmid replicon of the same contig 127. The presence of mcr-9 on a plasmid replicon facilitates its dissemination globally and across Enterobacteriaceae 127. It has thereafter been identified in more clinical settings 128–135 and in food-producing animals 136, 137. Among the 355 genomes, 65 were associated with a plasmid replicon, which may facilitate their dissemination globally 127. Mcr-9 cassette is usually located either on a IncHI2 and/or IncHI2A replicon or chromosomally encoded 127, 133, 138, 139. It is widely distributed within Enterobacteriaceae, particularly within Enterobacter sp. 129, 130, 132, 133, 135, Citrobacter telavivum 134, Salmonella sp. 127, 137, 138 and E. coli 136, 140.
Mcr-9 is always associated with wbuC, encoding a cupin fold metalloprotein, located downstream the gene 131, 135, 140. The amino acid sequence of mcr-9 closely resembles that of mcr-3 and mcr-7, at a 99.5% coverage; it has amino acid sequence identity of 64.5% to mcr-3.17 and 63% to mcr-7 127, 140. A three-dimensional structural model analysis of all nine mcr homologues based on the amino acid sequences of each showed that mcr-9 was closely related to mcr-3, mcr-4 and mcr-7 at a structural level 127. Further, all nine mcr-homologues had a high level of conservation for both domains (transmembrane domain and catalytic domain): the structural models showed the amino acids and structural elements were conserved in the C-terminal catalytic domain and the structural elements in the membrane-anchored N-terminal domain 127. A database search using mcr-9 as a template identified a chromosomal-encoded mcr-9-like gene within a Buttiauxella species isolate, a member of the Enterobacteriaceae 140. The identified mcr-like protein, MCR-BG, was isolated from B. gaviniae, and was found to have an 84% amino acid identity to the MCR-9 enzyme; MCR-BG, similar to mcr-9, has the wbuC gene located downstream 140.
Similar results were reported by Yuan et al. (2019) with a PEtN transferase isolated from B. brennerae. It had 86.83% protein sequence identity to mcr-9 at a 100% coverage 135. Further, the wbuC located adjacent the mcr-9 gene was found to be homologous to that of Buttiauxella with a 86% amino acid identity at a 98% coverage 135. This evidence suggests that mcr-9 may have originated from Buttiauxella species and disseminated to the rest of the Enterobacteriaceae family.
The dissemination of mcr-9 may have been aided by its association with IS elements flanking the cassette 133. The mcr-9 cassette has been found flanked with IS903 and IS903-like elements upstream as well as IS1R, IS26-like, and IS15DII located downstream in multiple reports 132, 133, 135, 140. The first report of the mcr-9 cassette encoded the wbuC gene, a TCS encoding qseC and qseB genes, ΔIS1R and a remnant of ΔsilR, encoding a transcriptional regulatory protein, all of which are located downstream of the mcr-9 gene. The mcr-9 cassette is thereafter flanked by intact IS903 elements 135. Though two copies of an intact IS element should be able to form a composite transposon or a circular intermediate, an inverse-PCR assay failed to detect any intermediates 135. However, the identification of IS903-like elements associated with mcr-9 genes suggests the acquisition of the cassette through an IS903 dependent mechanism. In some reports, the 3’region of mcr-9 is adjacent to copS (three copper resistance membrane-spanning proteins that includes rcnA, rcnR and a domain-containing DUF4942) 132, 139.
The mcr-9 protein as stated above has high levels of conservation within both PEtN transferase domains 127. Notwithstanding, multiple reports have shown mcr-9-harbouring isolates that are susceptible to colistin 127, 129, 130, with the colistin MIC value usually less than 2 mg/L 128. Kieffer et al. (2019b) 140 and Carroll et al. (2019) 127 performed induction experiments that conferred clinical resistance. However, the mechanism underlining each experiment remains unknown. Carroll et al. (2019) identified a conserved σ70 family-dependent -35 and -10 regions and an inverted repeat in the promoter of mcr-9 genes in the database. The authors suggest that the conserved DNA motif in the mcr-9 promoter is likely a recognition sequence for a transcription regulator, which might be needed for full expression of the mcr-9 gene 127. Kieffer et al. (2019b) 140 suggested the QseC-QseB TCS played a role in inducing mcr-9 expression to confer colistin resistance in the presence of sub-inhibitory concentration of colistin; however, multiple reports have failed to replicate these results 130, 134, 138, 139. In these studies, the mcr-9-harbouring isolates encoding the QseC-QseB TCS regulatory genes were fully susceptible to colistin 134, 138. Tyson et al. (2020) 138 suggests the induction and overexpression of mcr-9 may be context-dependent and be different in different strain backgrounds. The expression of mcr-9 in a pBAD vector conferred colistin resistance 141 and Kieffer et al. (2019b) 140 has shown that MCR-9 can mediate Lipid A modifications. However, the genes and molecules required to regulate mcr-9 expression in its own genomic environment remains unknown 130.
Mcr-10 variant
The mcr-10 gene was first identified in an Enterobacter roggenkampii clinical strain in China 141. It has a nucleotide sequence identity and amino acid sequence identity of 79.69% and 82.93% respectively to mcr-9 141. Therefore, like mcr-9, the mcr-10 gene shares a significant amino acid identity to the chromosomally encoded mcr-like PEtN transferase of Buttiauxella species 141. mcr-10 encodes a putative PEtN transferase that is made up of an N-terminal membrane-anchored domain and a C-terminal soluble catalytic domain, which have high levels of conservation to both MCR proteins and the MCR-B of various Buttiauxella species 141, 142. Xu et al. (2021) evaluated whether mcr-10 can mediate colistin resistance and compared its activity to that of mcr-1 and mcr-9. The study showed that the activity of mcr-10 against colistin is lower than that of mcr-9 but is able to mediate colistin resistance with an MIC value of 2.5 mg/L 142. This shows that mcr-10 is functional and thus aids the host to survive colistin selective pressure 142. Xu et al. (2021) again showed that the expression of mcr-10 is inducible under colistin resistance and further found an upregulation of phoP-phoQ TCS genes, producing a colistin MIC value of 8 mg/L. The authors suggested that mcr-10 might co-function with PhoPQ to mediate high levels of colistin resistance 142.
Mcr-10 has been identified on IncFIA and IncFIB plasmids 141–144 and adjacent to a XerC gene, which encodes a XerC type tyrosine recombinase found to mobilise adjacent genetic components such as antimicrobial resistance genes 141. Thus, mcr-10 would be presumed to have disseminated across Enterobacteriaceae family but has only been identified commonly in Enterobacter roggenkampii 141, 142, 145 and once in Cronobacter sakazakii 143. Wang et al. (2020a) and Xu et al. (2021) suggest that E. roggenkampii is an important reservoir of mcr-10.
In the initial identification of mcr-10, the gene was adjacent XerC and flanked by two intact IS903 elements which could form a composite transposon with the potential to mediate mobilisation of the mcr-10 gene 141. The analysis of the immediate genetic environment of the IS903 element found the absence of direct repeats that are generated during an insertion event; thus, the acquisition of the region bracketed by the IS903 elements was not due to an insertion event 141. The presence of an a ΔISEc36 downstream the mcr-10 gene was reported to make it impossible to identify the recombination sites that the XerC-type tyrosine recombinase recognizes 141. Thus, the acquisition events of mcr-10 into pMCR-10_09006 remains unknown 141. Mcr-10 has thereafter been 145 identified with various insertion sequences, including complete and truncated remnants of IS26 and ISKpn26, complete sequences of ISEc36 and IS1, a ΔISEcl1-like elements and recently, a new IS element designated ISCrsa1 143, 145. The identification of various IS elements downstream mcr-10-xerC suggests that this region might be a hotspot for insertions of MGEs 141, 142. The acquisition of mcr-10 is presumed to be through site-specific recombination; 143 however, evidence of mobilisation of mcr-10 is scarce.
Risk factors for acquiring mcr genes
Antimicrobials are commonly administered by farmers to pigs and poultry for prophylaxis, therapeutics, and growth promotion in livestock 146–148. Studies around the world have shown that the use of colistin as a growth promoter results in a high frequency of mcr-1 positive, colistin resistant Enterobacteriaceae (MPCRE) because of the high selective pressure in the veterinary environment. The selection pressure led to the acquisition and spread of mcr-1 genes 11, 146, 148–152. The presence of mcr-1 in food animals had a serious potential to spread MPCRE to humans via foodborne transmission 11, 150, 151.
The pharmaceutical form of colistin is indicated for oral administration and is present in a powder or solution form 152. Studies have shown that the drug is poorly absorbed in the digestive tract after oral administration and can be excreted in high levels through the faeces 1, 150, 152. This results in the presence of colistin or its active metabolites in the environment alongside the MPCRE isolates from the animal faeces 150. Xia et al. (2019) 153 measured the concentration of colistin in animal feeds and fresh manure in five swine farms and found that the average concentration of colistin in animal feed on farm 4 was 60 mg/kg, which was scientifically higher than other farms. The fresh manure samples collected on farm 4 had the highest concentration of 17,383 µg/kg. Other farms had colistin concentrations in their fresh manures ranging from 140 µg/kg in farms 1, 2 and 5, to 7,529 µg/kg in farm 3 153. The researchers also discovered a strong relationship between colistin concentration and the relative abundance of mcr-1 genes in fresh manure 153. The use of this manure as a fertilizer in agriculture or feeding farmed fish can result in the contamination of agriculture and, in some cases, the aquatic environment, polluting rivers and lakes 150, 154.
Luo et al. (2017) cultured an mcr-1 producing Raoultella ornithinolytica, a member of the Enterobacteriaceae family commonly found in soil, aquatic, and botanical environments, in retail vegetables in China. They emphasized that using animal excrements contaminated with colistin and/or MPCRE can cause contamination and spread of MPCRE to fresh vegetables. 154. There is, however, no epidemiological data on the contamination of rivers and lakes by MPCRE-contaminated animal excrements and thus, no evidence of the spread of MPCRE to the aquaculture 150. However, the presence of mcr-1-producing E. coli has been reported in duck and fish samples in China. Shen et al. (2019) 155 found a potential spread of mcr-1 producing E. coli (MPEC) from aquaculture supply chain to humans through aquatic food 150, 155. Other examples of the spread of MPCRE into the environment includes the identification and isolation of MPCRE in blowflies (Chrysomya sp.) 156 and black kites 54. Both cases are of public concern because of the potential transmission into human communities.
Yang et al. (2019a) 156 showed that blowflies may serve as an environmental reservoir and vector of MPCRE between animals, humans, the environment, and waste (landfills and sewage water) 156. The presence of MPCRE in Black kites in Russia highlights the spread of mcr-1 genes into both wildlife and the environment. Tarabai et al. (2019) 54 hypothesized that the acquisition of MPCRE was either through contact with the Biysk Municipal landfill or through their food, which was commonly found along the Biya River near their nest. Black kites can thereafter spread MPCRE along their long migratory pathways 54. The mcr-1 gene is commonly found on transferable plasmids, and thus the presence of mcr-1 producing isolates in the microbiota of insects and wildlife increases the possibility of horizontal transfer of mcr-1 genes alongside other resistance genes within the microbiota. This increases the environmental gene pool and the spread of multi-drug resistant (MDR) pathogens within insects and wildlife 156.
There are therefore multiple reservoirs highlighted above with the potential of spreading towards humans because of the close association between humans and the environment. A study in Vietnam showed that the farmers that are in contact with animals under colistin selection pressure have a high risk of being colonized by MPCRE 151. Trung et al. (2017) 151 found that chickens associated with colistin administration had a high carriage of mcr-1 producing isolates and the colonization of humans was through exposure to the chickens. The chicken faecal samples had a high prevalence of mcr-1 (59.4%) and zoonotic transmission prevalence was 34.7% in farmers. Another possible route of transmission of MPCRE from animals under colistin pressure was foodborne transmission 11, 157, 158. Monte et al. (2017a) found that chicken meat was acting as a reservoir of MPEC. A study performed by Shen et al. (2018) 150 found a positive correlation between MPEC carriage in the human normal flora and the consumption of farm animals, which included pork and sheep. The correlation analysis also proposed a possible transmission of MPEC from aquatic food to humans 150. This data is a single example that highlights the presence of MPCRE in food-producing animals and the transmission route of MPCRE from colistin-exposed animals to humans, which impacts public health care.
This therefore led to the ban on colistin use for growth promotion and disease prevention in the food industry to preserve their effectiveness in human medicine as their use contributes to the rising threat of antibiotic resistance 159. Randall et al. (2018) 160 also demonstrated that discontinuing colistin in farming can result in the elimination of mcr-1. This longitudinal study evaluated the presence of mcr-1 in pig faeces and slurry at two time points 160. At the beginning of the study, the majority of the pigs were colonized by MPEC. Twenty months after cessation of colistin use and implementation of hygiene and other measures, MPEC was not detected in samples. These results thus highlight that the use of colistin is the driving force behind the spread of colistin resistance genes and its stability in the population 160. The integration of mcr-1, however, into conjugative plasmids encoding other resistance genes allows for the use of other antibiotics such as quinolones and extended spectrum cephalosporins to simultaneously co-select for mcr-1, allowing for its stability and dissemination 146, 161, 162. This, thus, creates severe challenges for controlling the selection and subsequent transmission of mcr-1 genes 163.
After the first detection of mcr-1 by Liu et al. (2016), mcr-1 genes have been detected in animals, the environment, and humans. Initially, the detection of mcr-1 genes in humans was associated with the risk factors mentioned above: contact with farm animals, ingestion of contaminated farm food, or fresh vegetables. Epidemiology studies that identified mcr-1 genes in the clinical settings were accompanied with a questionnaire to identify the epidemiological exposure. In countries such as Finland, where the use of colistin is restricted for rare clinical indications, the first detection of the mcr-1 gene was identified in a healthy male with previous history of travelling abroad, 6 months before faecal sampling 164. Traveling to other countries has been shown to increase the risk of acquiring mcr-1 positive isolates, particularly in patients who consumed meat while traveling abroad 164, 165. The use of antibiotics prior to hospitalization or infection has been seen as an important risk factor for a multidrug resistant organism (MDRO) infection 165. An epidemiological and clinical study performed by Wang et al. (2017) 166 discovered that MPEC can acquire other resistance genes, and thus previous use of antibiotics such as carbapenems and fluroquinolones was associated with an increased risk of infection by MPEC 166. Thus, the administration of antibiotics in clinics simultaneously promotes the co-selection and the preservation of colistin resistance genes 167.
An interesting possible transmission of MPEC is between companion animals and humans. A worker at a pet shop with no prior antibiotic use or travelling abroad was identified carrying an MPEC 168. A faecal screening of the pets residing where the man worked identified 4 dogs and 1 cat with an mcr-1 positive isolate. Zhang et al. (2016) 168 suggested companion animals as a possible reservoir of colistin-resistant E. coli, facilitating the spread of colistin resistance genes within the community.
Current and future perspectives on treating mcr-mediated colistin resistance
Clustered regularly interspaced short palindromic repeats (CRISPR)
Clustered regularly interspaced short palindromic repeats (CRISPR) has been exploited and developed within molecular biology as a site-specific tool for genetic engineering 169, and in regards to resistance, as a tool to deliver a programmable DNA nuclease (CRISPR-Cas9) into an MDR-pathogen to eliminate the resistance gene or plasmid 170. Dong et al. (2019) 170 developed a conjugative CRISPR-Cas9 system that aimed to remove plasmids harbouring the mcr-1 gene from bacteria. The system targeted the mcr-1 gene and the authors found that the formation of DSBs during the elimination of the resistance gene resulted in the elimination of the whole plasmid in recipient cells 170. This thereafter sensitized the recipient cells to colistin. Dong et al. (2019) 170 further found that the cells are thereafter immune against the mcr-1 gene. Wang et al. (2019a) 171 used the same concept but developed the tool to remove both mcr-1-harbouring plasmids and MDR plasmids present in recipient cells using partial sequences of the targeted plasmids. He et al. (2021) 172 used the tool to eliminate both chromosomal and plasmid-borne mcr-1 genes in E. coli. The CRISPR-Cas9 system was designed to target ISApl1, which eliminated mcr-1 harbouring plasmids, and in the case of chromosomally encoded mcr-1, cell death was seen. Similar results were achieved 170, where recipient cells further acquired immunity against the acquisition of the exogenous mcr-1 containing plasmid 172. These studies show that CRISPR-Cas9 system is an efficient tool for plasmid curing and to sensitise clinical isolates to antibiotics in vitro 170, 171. This tool can be optimized for therapeutic application for the elimination of antibiotic genes from resistance reservoirs such as animal guts with prior exposure to antibiotics, the human microbial flora or the bacteria in natural settings such as wastewater, farms or hospital settings 170.
The CRISPR-Cas9 system is however made up of highly anionic nucleic acids and proteins that cannot penetrate the cell membrane into cells and thus requires a delivery vehicle. Sun et al. (2017c) 173 explored Cathelicidins, which are short cationic antimicrobial peptides, as a plasmid delivery system. The study evaluated the use of BMAP-27 to increase the efficiency of plasmid transfer of the CRISPR-Cas9 system and found that pCas::mcr exhibited better, efficient, and specific antimicrobial effects with the help of BMAP-27 173. The CRISPR-Cas9 system is a powerful tool for genome editing, its development into targeting multiple genes with one use 171, could aid in the removal of mcr variants in resistance reservoirs and in clinical settings to restore colistin activity.
In the meantime, there are multiple studies evaluating existing antibiotics for activity against mcr-producing isolates and synergy combinations.
Novel antibiotics
There are multiple novel antibiotics that have been found to have activity against mcr-1-producing isolates, including eravacycline 6, 174 and plazomicin 175. Eravacycline is a broad-spectrum synthetic tetracycline that has been proven to be effective against clinically important MDR pathogens, including both Gram-negative and gram-positive bacteria 174. Fyfe et al. (2016) 6 found that eravacycline had a bactericidal effect against mcr-producing isolates from Enterobacteriaceae. The study further showed that although the overexpression of mcr-1 increased colistin MIC values by 64-folds, there was no effect on eravacycline susceptibility 6. Eravacycline activity was thereafter evaluated against 336 isolates collected from hospitals across China and was found to exhibit a good efficacy against all strains 174. These isolates included Extended spectrum β-lactamase (ESβL) and carbapenemase-producing Enterobacteriaceae, mcr-producing Enterobacteriaceae and A. baumannii, vancomycin-resistant Enterobacteriaceae, β-lactamase producing Haemophilus influenzae, penicillin-resistant Streptococcus pneumoniae and lastly, methicillin-resistant Staphylococcus aureus (MRSA), thus showing a positive potential to treat current drug-resistant bacterial infections 174. This, however, also puts eravacycline at risk of being overused.
Plazomicin is a novel broad-spectrum semi-synthetic aminoglycoside that was derived from Sisomicin. It has been shown to have activity against a broad spectrum of MDR pathogens, including ESβL- and carbapenemase-producing isolates and fluroquinolone-resistant isolates175. Denervaud-Tendon et al. (2017) 175 evaluated the bactericidal activity of plazomicin against colistin-resistant Enterobacterial strains, which harboured different colistin resistance mechanisms i.e., chromosomal-encoded, mcr-1-expressing, and intrinsically resistant strains. Plazomicin activity was further compared with other aminoglycoside antibiotics viz., amikacin, gentamicin, and tobramycin. Plazomicin displayed potent activity against the clinical colistin-resistant Enterobacterial strains irrespective of their resistance mechanisms. However, those that were intrinsically resistant (Serratia, Proteus, Morganella and Hafnia) had a higher MIC value 175.
Denervaud-Tendon et al. (2017) also found that amongst all aminoglycoside antibiotics tested, plazomicin was the most potent. They lastly also found that plazomicin’s activity is restricted by aminoglycoside resistance mechanisms such as the 16S rRNA methylase- encoding gene, which resulted in an elevated plazomicin MIC value (>128 mg/L). Plazomicin, however, like other aminoglycosides, has a rapid bactericidal activity with favourable chemical and pharmacokinetics properties and thus could be incorporated into therapeutic treatments against mcr-producing bacterial infections 175.
Artilysin ® Art-175 is a novel engineered enzyme-based experimental therapeutic derived from endolysins produced by lytic bacteriophages. These endolysins degrade the bacterial cell wall resulting in cell lysis, which occurs at the end of the bacteriophage infection cycle 176. Artilysin ® Art-175 is made up of the outer membrane destabilizing peptide, which is fused to an endolysin, it targets the anionic lipopolysaccharide molecules of the outer membrane of Gram-negatives. Schirmeier et al. (2018) 177 evaluated the potential of Art-175 against pan-drug resistant isolates, including colistin-resistant MPEC and further determined the potential of cross-resistance between colistin and Art-175. Art-175 was highly bactericidal against the tested isolates, which were more susceptible to Art-175 than the colistin-susceptible isolates 177. Hence, there is no cross-resistance between the two peptides, highlighting Art-175 as a potent solution against mcr-producing isolates 177.
Antimicrobial peptides
Bacteriocins are ribosome-synthesized antimicrobial peptides (AMPs) produced by both Gram-positive and Gram-negative bacteria. Studies have shown that bacteriocins produced by lactic acid bacteria (LAB) are cationic peptides with bactericidal activity that act on the cytoplasmic membrane of susceptible microorganisms 178. The LAB bacteriocins create pores in the membrane, resulting in intracellular damage. Al Atya et al. (2016) 4 evaluated the combinations of nisin and enterocin DD14 LAB bacteriocins with colistin to eradicate colistin-resistant E. coli strains in either planktonic states or in biofilm. Whilst colistin and the bacteriocins were ineffective as monotherapy against both planktons and biofilms, colistin-nisin and colistin-enterocin DD14 combination were able to reduce the number of colony-forming units (CFU) of strains in both planktonic and biofilm states. The triple combination of all three molecules, colistin-nisin-enterocin DD14, completely eradicated all E. coli strains, including colistin-resistant phenotype 4. LAB bacteriocins alone cannot penetrate the outer membrane. However, with the disruption of the LPS via colistin activity, the LAB bacteriocins may thereafter act upon the cytoplasmic membrane 4. Al Atya et al. (2016) 4 therefore highlights LAB bacteriocins as a potential novel adjunctive treatment for colistin-resistant MPEC infections.
There have been multiple reports of AMPs that have been developed for treating MDR pathogens, which have a similar mode of action as colistin 179. Van der Weider et al. (2019) evaluated the antimicrobial activity of two novel AMPs, AA139 and SET-MM3. AA139 originates from a marine lugworm, Arenicola marina, AMP arenicin-3. SET-MM3 is a synthetic tetra-branched peptide linked by a lysine core 179. The antimicrobial activity of these AMPs was evaluated against a collection of clinically and genotypically diverse K. pneumoniae with different antimicrobial susceptibility profiles. Both AMPs, AA139 and SET-M33, further showed a concentration-dependent bactericidal effect across all isolates, meaning that the susceptibility profile of both AMPs was unaffected by the resistance profile and colistin susceptibility of the isolates 179. The AMPs were effective against the colistin-resistant strains, and no cross-resistance was observed between the AMPs and colistin. Thus the antimicrobial activity of AA139 and SET-M33 should further be investigated across other Gram-negatives to evaluate its spectrum of bactericidal activity 179.
Cross-resistance between several human cationic AMPs (CAMPs) and polymyxins has been previously reported in PEtN transferase-producing Haemophilus ducreyi and Campylobacter jeyuni but rarely in Enterobacteriaceae. These CAMPs are part of the intrinsic human immune system and act against Gram-negative bacteria by disrupting the outer and inner membrane through electrostatic interaction, similar to colistin’s mechanism of action180. Dobias et al. (2017) 180 selected a few widely distributed CAMPs in humans, LL-37, α-defensin 5 (HD5) and β-defensin 3 (HDB3) and evaluated whether cross-resistance between defensin 5 (HD5) and β-defensin 3 (HDB3) and evaluated whether cross-resistance between CAMPs and polymyxin would be observed in MPEC. Susceptibility of E. coli to the different CAMPs selected varied depending on the CAMP molecule rather than the entire CAMP family. It further showed that the production of MCR proteins did not confer cross-resistance with the human CAMPs selected 180.
However, the CAMPs activity studied by Dobias et al. (2017), 180 compared to other studies 181, shows that factors such as bacterial species, level of resistance, and colistin resistance mechanisms may affect the bactericidal levels of CAMPs 180, 181. Therefore, although mcr production did not affect the bactericidal activity of CAMPs against E. coli, it needs to be evaluated across Enterobacteriaceae and other Gram-negative bacteria to see how different colistin resistance mechanisms affect the CAMPs’ bactericidal effect. Guachalla et al. (2018) 182 further evaluated the effectiveness of other derivates of the human immune system against a clinical MPEC strain. This includes humanized monoclonal antibodies that have been previously shown to target the LPS O25b antigen that is associated with E. coli ST131-H30 182. The expression of mcr-1 does not affect the binding of ASn-4 to the LPS’s O-antigen and the integration of the complement membrane-attack complex (MAC) into the LPS-modified outer membrane 182. This means the expression of mcr-1 in E. coli ST131-H30 does not affect the functionality of the human immune system, nor does it not aid the strain via immune evasion. The authors thereafter suggested the use of antibodies targeting the LPS’s O-antigen as an alternative strategy to combatting MDR infections against mcr-1 positive isolates 182.
Other novel compounds produced to manage mcr-producing isolates are antisense agents with antimicrobial properties; this include phosphorodiamidate morpholino oligomers (PPMOs) and peptide nucleic acids (PNAs) 183. These antisense molecules are designed to target mRNA, prevent translation, and restore antimicrobial sensitivity. Daly et al. (2017) 184 designed peptide-conjugated PPMOs targeted to mcr-1 mRNA (MCR-1 PPMO) of clinical mcr-1-positive E. coli strains and showed that these molecules were able to make the strains re-sensitive to polymyxins by MCR-1 inhibition 184. Similar results were achieved by Nezhadi et al. (2019) 183 with PNAs, designed for inhibition of mcr-1 translation. They 183 showed that the introduction of PNA resulted in a 95% reduction of mcr-1 expression, measured by RT-PCR, highlighting the efficacy of PNAs 183. Daly et al. (2017) 184 evaluated the antisense approach in a sepsis model using mice and found that a combined therapy of MCR-1 PPMO with colistin reduced morbidity and bacterial burden in the spleen at 24hr. Thus, antisense agents may be effective therapeutics, either alone by targeting an essential gene resulting in cell death, 184 or in combination with colistin 183, 184 to treat mcr-1-producing isolates. However, these molecules have varying toxicity effects and more in-vivo research is required prior to clinical application 183.
Natural compounds
Other novel compounds that have been reported to restore susceptibility of polymyxins in mcr-1 producing isolates includes the novel MCR-1 inhibitor, Osthole (7-methoxy-8-(3-methyl-2-buteryl) coumarin. Osthole (OST) is a natural compound derived from the dried root and rhizome of Cnidium monnieri 185. Zhou et al. (2019) 185 evaluated the effects of OST on the inhibition of MCR-1 enzyme and the mechanisms behind the inhibition. The colistin-OST combination had a synergistic effect as a mouse-thigh-infection model showed that the combination exhibits a bactericidal activity against mcr-1-producing Enterobacteriaceae, significantly reducing the bacterial load in the thighs following subcutaneous administration 185. Using a molecular dynamic stimulation, Zhao et al. (2019) 185 found that OST could localize in the binding pocket (residue 330-350) of MCR-1, blocking the substrate and reducing MCR-1’s biological activity. OST therefore inhibits MCR-1 in Enterobacteriaceae, making colistin potent 185. Due to the conservation of MCR proteins, OST may have the same inhibition efficiency across the other MCR variants.
Natural/organic compounds identified with a synergistic effect on colistin include Honokiol 186, Isoalantolactone (IAL) 187, Calycosin 188 and Eugenol 189. Honokiol was derived from the dry skin and bark of Magnolia officinalis 186, IAL is the main sesquiterpene lactone from Radix inulae and other plants 187, Calycosin is a flavonoid from a direct root extract of the traditional Chinese medicinal herb, Radix astragali.188 Lastly, Eugenol is a phenylpropanoid, an essential oil isolated from plants 190. Each of these compounds, when combined with colistin, decreased colistin MICs and increased colistin’s bactericidal activity against colistin-resistant isolates. Eugenol decreased the expression of mcr-1 189 and honokiol bound directly to the active site of MCR-1, inhibiting its activity 186. Guo et al. (2020) 186 and Liu et al. (2020) 188 showed that the natural compounds, honokiol and calycosin, in combinations with colistin, respectively reduced the load of bacteria and improved viability in animal models.
Efflux pump inhibitors
Efflux pumps play a role in reducing colistin susceptibility in Enterobacteriaceae 10. Baron et al. (2018) 191 found that an efflux pump inhibitor, carbonyl cyanide 3-chlorophenylhydrazone (CCCP), was a good alternative to reverse colistin resistance in colistin-resistant isolates irrespective of their molecular resistance mechanisms. The study shows that CCCP was able to restore colistin activity across a diverse set of strains carrying different colistin resistance mechanisms (mcr-1, pmrAB, mgrB, etc) 191, 192. In mcr-1 producing isolates, it was seen that colistin-CCCP combination inhibited the transcription of the mcr-1 gene. The mechanism behind this observation, however, is unknown 191.
FDA-approved drugs
The food and drug administration (FDA) has a library of FDA-approved drugs that can be screened for bactericidal activity on mcr-producing Enterobacteriaceae isolates (Prestwick Chemical, Illkirch-Graffenstuden, France). Multiple compounds such as pentamidine, zidovudine 193, sulphonamide compounds 193, a polymyxin derivative, NAB739, 194 and compound PFK-185, 194 have been identified during a library search.
Zidovudine is a nucleoside reverse transcriptase inhibitor. In the 1980s, it was used as an anticancer drug 195. In 1986, its antibacterial effect was revealed 196 and in 1987, it was used as the first antiretroviral for treatment against HIV infections 195, 197. Peyclit et al. (2018) 197 investigated the antibacterial activity of Zidovudine against a large number of characterised MDR Enterobacteriaceae strains isolated from different geographical areas, including carbapenem- and colistin-resistant isolates; Zidovudine was effective against Enterobacteriaceae. Despite their antimicrobial susceptibility profile, the MIC values ranged between 0.05-1.67 µg/mL 197.
Another antiretroviral drug that is used to treat HIV/AIDS but also has bactericidal activity against Gram-negative bacteria is Azidothymidine (AZT). Hu et al. (2019) 198 shows that AZT, in combination with colistin, was able to eradicate MDR Enterobacteriaceae strains with different antimicrobial susceptibility profiles and enhance the activity of colistin. The combined therapy was also effective therapeutically in a murine peritoneal infection against NDM-1-producing K. pneumoniae and MPEC 198.
Further, Pentamidine is an antiprotozoal drug that Stokes et al. (2017) 199 identified as an effective antibiotic adjuvant producing synergistic combination activity against a wide range of Gram-negative bacteria. The adjuvant was identified during a library screening of non-lethal, outer membrane-active compounds 199. The study shows that Pentamidine directly associates with the outer membrane by inhibiting the oligosaccharide (OS) core biosynthesis, releasing LPS from the outer membrane 199. Stokes et al. (2017) 199 further showed that Pentamidine has synergistic activity with hydrophobic antibiotics such as rifampicin, novobiocin, and erythromycin but not with hydrophilic and low-molecular weight antibiotics 199. Although the study does not evaluate synergistic combinations with Pentamidine, Stokes et al. (2017) 199 concluded that pentamidine can be used as an antibiotic adjuvant for treating colistin-resistant infections.
Barker et al. (2017) 200 screened a diverse cohort of adjuvants to identify a compound that could both sensitise colistin-resistant and hypersensitise colistin-susceptible bacteria to colistin, to potentially lower the effective dosage of colistin, reducing its toxicity. The study identified three compounds that were potent modulators of colistin resistance and were able to significantly reduce MIC values in both plasmid-and chromosomal-mediated resistance mechanisms, and further hypersensitize colistin-susceptible isolates 200
Another compound identified in the FDA-approved library with synergistic activity with colistin is sulfadiazine (SDI), a sulphonamide compound. Okdah et al. (2018) 193 evaluated the potential of different sulphonamide compounds with potential synergistic and bactericidal activity in combination with colistin; SDI had the highest synergistic effect. The combination of SDI-colistin was effective, independent of colistin resistance mechanism, across the broad range of MDR bacterium strains tested 193.
Zhang et al. (2019) 194 identified an antitumor drug, PFK-185 and its analogs, PFK-015 and 3PO, during a screening of the clinical compound library. These compounds can exert synergise with colistin against colistin-resistant Enterobacteriaceae despite mcr expression and the antimicrobial susceptibility profile 194. Zhang et al. (2019) 194 found that PFK-185 had no effect on cellular morphology when used alone. However, when in combination with colistin, it enhanced the bacterium-killing effect of colistin, increasing the survival rate to 60%. The combined colistin-PFK-158 therapy had the most significant bactericidal activity, with the most significant reductions in the bacterial burdens post-in vivo experiments and during time-kills studies. 194.
The last compound reported from the FDA-approved library is a novel polymyxin derivative, NAB739, which carries only three of the five amino groups of colistin, each placed in a strategic position 201. Tyrrell et al. (2019) 201 showed that NAB739 sensitizes polymyxin-resistant strains to rifampicin and a combination of the two was synergistic against ten of the eleven colistin-resistant strains. Tyrrell et al. (2019) 201 further showed that NAB739 was also synergistic with meropenem and retapamulin. As well, NAB739 in combination with other compounds has significant bactericidal activity against polymyxin-resistant isolates and compared to polymyxins, it has a better tolerability and efficacy 201. Thus, NAB739 combination therapies may replace polymyxin for treating MDR pathogens 201.
These studies 197, 199 however, show that old antimicrobials listed within the FDA-approved library may be re-introduced against bacteria that are resistant to multiple antibacterials in current use 197.
Combination therapy
There are multiple compounds, either antibiotics or adjuvants, which are synergistic with colistin, thus reducing the dosage of colistin required for treatment, minimizing its toxicity, overcoming colistin resistance, and retaining maximal therapeutic efficacy 198, 200. Colistin activity has been restored when it is combined with antibiotics such as rifampicin 202, 203, rifabutin, minocycline 203, amikacin 204, tigecycline 205 and clarithromycin 206. In all these reports, the antibiotics were ineffective as monotherapy against MPCRE isolates. However, in combination with colistin, they resulted in significant bactericidal activity. Yu et al. (2019) 203 found that colistin in combination with rifampicin, rifabutin or minocycline was able to eradicate XDR, NDM- and mcr– co-producing E. coli in-vitro and in mouse models. Other combinations that were confirmed effective against mcr-1-producing colistin-resistant E. coli using an animal model include tigecycline-colistin 205, clarithromycin-colistin 206 and a triple combination of colistin-rifampin-azithromycin discovered by Li et al. (2018b) 202. Another triple combination therapy that was discovered was colistin-aztreonam-amikacin, which was effective against both carbapenemase-producing and mcr -1-producing Enterobacteriaceae isolates 207.
Lastly, a combination therapy that was effective against both carbapenem- and colistin-resistant Enterobacteriaceae isolates was the β-lactam-β-lactamase inhibitor, imipenem-relebactam, combination 208. This combined therapy was potent against colistin-resistant carbapenemase-producing isolates, making it a potential agent against carbapenemase-producing Enterobacteriaceae; particularly, those that are colistin resistant (irrespective of the mechanism of resistance) 208.
Conclusion
Herein, we show that mcr genes are commonly identified in E. coli, K. pneumoniae, and Salmonella sp., with clones within these strains being disseminated globally within animals, food, the environment, and humans. Mcr genes were initially identified in animal samples, specifically livestock animals that were treated with colistin as a therapeutic against Gram-negative bacterial infections or as a growth promoter. Mcr has spread to the environment through contaminated but untreated animal faeces used as manure and through contaminated food-producing-animals’ products on markets. Contact with these colistin-treated livestock or their faeces by humans was an identified transmission route. Human specimens currently have the highest reports of mcr genes due to the multiple routes of transmission from livestock, the environment and from food, and the use of colistin as a therapeutic against carbapenem-resistant Enterobacteriaceae infections. The presence of clones such as E. coli ST744 or ST101 in all sources, including animals, the environment, food, and humans, highlights these transmission routes. This makes mcr a concerning antibiotic resistance gene of great interest and priority.
Further, mcr genes are commonly associated with mobile genetic elements. ISApl1 elements have facilitated the horizontal transfer of mcr-1 within cassettes of composite transposons. They have been also associated with IncX4 plasmids, facilitating their horizontal and vertical transfer across species, genera and families.
Although the mcr variants have different possible progenitors viz., Moracella sp., Aeromonas sp., and Shewanella sp. for mcr-1, mcr-3 and mcr-4, respectively, the MCR proteins are very well conserved. Each of the 10 MCR proteins encodes a two-domain integral membrane protein with a C-terminal periplasmic domain and an N-terminal 5’-helix transmembrane domain, each MCR protein further encodes the five conserved residues i.e., E248, T286, H389, D458, and H359, located within the active site. PEtN transferase activity was found for each MCR protein, and each was able to mediate colistin resistance, although mcr-9 expression requires a specific genetic environment to regulate its expression. The conservation seen within MCR proteins, allows for the use of similar therapies for the management of MCRPE isolates, specifically in the cases of identified MCR-1 inhibitors. Research on novel therapeutics is well-summarised in this review, with techniques such as CRISPR-Cas9, peptide nucleic acids, and antimicrobial peptides, which eliminate mcr genes within the host. Other novel therapeutics were identified by reviving old FDA-approved drugs. Some were effective against mcr-producing isolates or were synergistic with colistin.
These therapeutics, however, need to be further evaluated for their toxicity in humans. This will aid in alleviating the threat imposed by MCRPE in the public health sector
Data Availability
All data are attached as supplementary files
Funding
This work was funded by a grant from the National Health Laboratory Service (NHLS) given to Dr. John Osei Sekyere under grant number GRANT004 94809 (reference number PR2010486).
Transparency declaration
None
Acknowledgements
None
References
- 1.↵
- 2.↵
- 3.↵
- 4.↵
- 5.↵
- 6.↵
- 7.↵
- 8.
- 9.↵
- 10.↵
- 11.↵
- 12.↵
- 13.↵
- 14.↵
- 15.↵
- 16.↵
- 17.↵
- 18.↵
- 19.↵
- 20.↵
- 21.↵
- 22.↵
- 23.↵
- 24.↵
- 25.
- 26.↵
- 27.↵
- 28.↵
- 29.↵
- 30.↵
- 31.↵
- 32.↵
- 33.↵
- 34.↵
- 35.↵
- 36.↵
- 37.↵
- 38.↵
- 39.↵
- 40.↵
- 41.↵
- 42.↵
- 43.↵
- 44.↵
- 45.↵
- 46.↵
- 47.↵
- 48.↵
- 49.↵
- 50.↵
- 51.↵
- 52.↵
- 53.↵
- 54.↵
- 55.↵
- 56.↵
- 57.↵
- 58.↵
- 59.↵
- 60.↵
- 61.↵
- 62.↵
- 63.↵
- 64.↵
- 65.
- 66.↵
- 67.↵
- 68.↵
- 69.↵
- 70.↵
- 71.↵
- 72.↵
- 73.↵
- 74.↵
- 75.↵
- 76.↵
- 77.↵
- 78.↵
- 79.↵
- 80.↵
- 81.↵
- 82.↵
- 83.↵
- 84.↵
- 85.↵
- 86.↵
- 87.↵
- 88.↵
- 89.↵
- 90.↵
- 91.↵
- 92.↵
- 93.↵
- 94.↵
- 95.↵
- 96.↵
- 97.↵
- 98.↵
- 99.↵
- 100.↵
- 101.↵
- 102.↵
- 103.↵
- 104.↵
- 105.↵
- 106.↵
- 107.↵
- 108.↵
- 109.↵
- 110.↵
- 111.↵
- 112.↵
- 113.↵
- 114.↵
- 115.↵
- 116.↵
- 117.↵
- 118.↵
- 119.↵
- 120.↵
- 121.↵
- 122.↵
- 123.↵
- 124.
- 125.
- 126.↵
- 127.↵
- 128.↵
- 129.↵
- 130.↵
- 131.↵
- 132.↵
- 133.↵
- 134.↵
- 135.↵
- 136.↵
- 137.↵
- 138.↵
- 139.↵
- 140.↵
- 141.↵
- 142.↵
- 143.↵
- 144.↵
- 145.↵
- 146.↵
- 147.
- 148.↵
- 149.
- 150.↵
- 151.↵
- 152.↵
- 153.↵
- 154.↵
- 155.↵
- 156.↵
- 157.↵
- 158.↵
- 159.↵
- 160.↵
- 161.↵
- 162.↵
- 163.↵
- 164.↵
- 165.↵
- 166.↵
- 167.↵
- 168.↵
- 169.↵
- 170.↵
- 171.↵
- 172.↵
- 173.↵
- 174.↵
- 175.↵
- 176.↵
- 177.↵
- 178.↵
- 179.↵
- 180.↵
- 181.↵
- 182.↵
- 183.↵
- 184.↵
- 185.↵
- 186.↵
- 187.↵
- 188.↵
- 189.↵
- 190.↵
- 191.↵
- 192.↵
- 193.↵
- 194.↵
- 195.↵
- 196.↵
- 197.↵
- 198.↵
- 199.↵
- 200.↵
- 201.↵
- 202.↵
- 203.↵
- 204.↵
- 205.↵
- 206.↵
- 207.↵
- 208.↵
- 209.↵
- 210.↵




