
Abstract
Background and objectives Low socioeconomic status (SES) is consistently associated with chronic stress, poor health, and premature death in high income countries (HICs). However, the degree to which SES gradients in health are universal—or even steeper under contemporary, post-industrial conditions—remains poorly understood.
Methodology We quantified material wealth and several health outcomes among a population of traditional pastoralists, the Turkana of northwest Kenya, who are currently transitioning toward a more urban, market-integrated lifestyle. We assessed whether SES associations with health differed in subsistence-level versus urban contexts. We also explored the causes and consequences of SES-health associations by measuring serum cortisol, potential sociobehavioral mediators in early life and adulthood, and adult reproductive success (number of surviving offspring).
Results High SES predicts better self-reported health and more offspring in traditional pastoralist Turkana, but worse cardiometabolic health and fewer offspring in urban Turkana. We do not find robust evidence for either direct biological mediators (i.e., cortisol) or indirect sociobehavioral mediators (e.g., adult diet or health behaviors, early life experiences) of SES-health relationships in either context.
Conclusions and implications While social gradients in health are common in non-human primates, small-scale human societies, and HICs, we find that relationships between wealth and health can vary dramatically within a single population. Our findings emphasize that under different economic and societal circumstances, social status effects on health may manifest in very different ways.
Lay summary High socioeconomic status predicts better health and more offspring in traditional Turkana pastoralists, but worse health and fewer offspring in individuals of the same group living in urban centers. Together, our work shows that under different economic and societal circumstances, wealth effects on health may manifest in very different ways.
Introduction
Social gradients in health exist in almost every human population that has been studied. In the United States, individuals in the lowest socioeconomic class (defined by income) are at greater risk for major health issues such as heart disease, cancer, and diabetes, and are predicted to die over a decade earlier than individuals in the highest socioeconomic class [1–3]. These socioeconomic status (SES) gradients in disease risk and survival are to some degree explained by differences in health care, health habits, and access to resources that are also socially stratified [4]. However, studies in animal models, where such confounds can be avoided, support the hypothesis that some portion of the social status-health relationship is driven by direct and causal effects of social status on physiology, largely mediated by chronic stress and altered HPA axis function in low status individuals [5–7].
While there is overwhelming support for a consistent relationship between wealth-based measures of SES and human health, most evidence to date comes from studies of high-income countries (HICs) [8,9]. This limited scope undermines our ability to understand whether SES-health relationships are consistent across contexts, or instead vary as a function of resource availability and distribution, the nature of social relationships and hierarchies, or other socioecological features of a population (as has been shown for other species [10–12]). In particular, it has been hypothesized that the steep social gradients in health observed in “Western” HICs are recent byproducts of environmental changes precipitated by urbanization, globalized markets, capitalism, and other modern advancements [13,14]. In other words, while social gradients in health have deep roots in primate and human evolution [6,7], the nature and magnitude of these gradients have potentially changed as a function of modern lifeways.
There are several potential explanations for why post-industrial conditions may exacerbate wealth/SES effects on health. First, relative to small-scale, subsistence-level groups such as hunter-gatherers, modern societies exhibit limited upward mobility and reduced kin support, as well as deep structural racism and violence which may intensify stress in the lowest socioeconomic strata [13,15,16]. Second, modern societies also exhibit a long list of socially stratified health care resources and health habits (e.g., obesogenic diet, drug and alcohol use) that were largely absent during pre-industrial periods [17–20]. Finally, epidemiological changes that go hand in hand with industrial transitions could alter the nature of SES-health relationships: most deaths in modern day HICs are attributed to non-communicable rather than infectious diseases, and these disease classes are likely differentially affected by wealth However, because there is a relatively limited literature examining SES effects on health outside of the industrialized setting (e.g., in pre-industrial societies or small-scale, subsistence-level groups), the degree to which urbanization and market-integration fundamentally change the strength or nature of SES-health relationships remains poorly understood [13,21–26].
To address this gap, we quantified the relationship between SES (operationalized as material wealth) and health in a small-scale, subsistence-level pastoralist population—the Turkana people of northwest Kenya. Pastoralists are often portrayed as egalitarian, largely because of their relatively equal and open access to natural resources, trading of livestock holdings, and resource sharing during times of hardship [27,28]. The social networks and relationships between herd owners are thought to be extremely strong and fundamental to the pastoralist way of life [28,29]. However, livestock holdings among pastoralists are also highly correlated across generations, and the intergenerational transmission of inequality is on par with or even greater than what is observed in the most unequal HICs [30]. Among the Turkana specifically, there is extreme variation in livestock holdings within and between generations, crossing several orders of magnitude and often fluctuating due to unpredictable events such as droughts, livestock disease outbreaks, or raiding from nearby groups [28]. This complex picture of high wealth inequality and variance paired with egalitarian practices makes it unclear to what degree we should expect SES to affect health outcomes in pastoralist societies like the Turkana. While previous work with pastoralist communities has examined effects of herd size (the primary source of material wealth) on nutrition and caloric intake [31–33], little work has tested the relationship between SES and health in traditional pastoralist societies [32,34,35].
Our study set out to explore SES-health connections in traditional Turkana, and to ask how these links strengthen or change when individuals transition to a more urban, market-integrated lifestyle. We were able to perform this comparison because cultural and economic changes paired with expansion of country-wide infrastructure has prompted many Turkana to move to densely populated cities over the last few decades. Turkana migrants to urban areas no longer practice pastoralism, work wage labor or market-interfacing jobs, and experience many other lifestyle changes. For example, urban-dwelling Turkana consume fewer traditional and more processed foods relative to pastoralist Turkana, which puts them at greater risk of cardiometabolic disease [36]. Moving to urban areas may also lead to increased psychosocial stress, reduced kin support, and changes in health habits and physical activity—all of which impact health and may vary with SES [7,37,38].
By collecting data on wealth and health from both traditional, pastoralist and urban, market-integrated Turkana, we were able to test whether lifestyle change alters SES effects on health within a single population. We also performed several follow up analyses to understand the causes and consequences of SES-health associations in both rural and urban contexts. First, to understand the putative fitness consequences of SES, which is essential for understanding how the behavioral and physiological correlates of social status evolve [39], we tested for SES effects on reproductive success (number of surviving offspring). Second, to understand the biological and behavioral mediators of SES-health connections, we measured 1) serum cortisol to assess the role of psychosocial stress and HPA axis function and 2) we interviewed study participants about their diet, health habits, and use of health care resources; we then asked whether SES predicted any of these potential mediators, and if so, we performed formal mediation analyses [40,41]. Finally, we were interested in understanding the role of another key dimension of the social environment—early life adversity—which has been shown in HICs to both affect later life health outcomes [6,42,43] and to set individuals on a course toward low SES in adulthood [43–45]. We therefore devoted effort toward systematically documenting variation in early life experiences and exploring their effects, which has rarely been attempted in small-scale, subsistence-level groups [46–48]. Taken together, our study provides a comprehensive picture of how, why, and under what circumstances social status affects health. We leverage the lifestyle gradient of the Turkana to directly address the impact of increasing urbanization and market-integration on this important relationship.
Methods
Overview of the study population and study methodology
The Turkana have resided in northwest Kenya since the early 18th century [49]; their homelands (Turkana county) are semi-arid and characterized by low annual rainfall, frequent droughts, and high year round temperatures [50]. The Turkana people are traditionally nomadic pastoralists, relying on dromedary camels, zebu cattle, fat tailed sheep, goats, and donkeys for subsistence [51]. Most herders keep livestock from all species, though they may specialize to some degree [29]. As a result of their subsistence strategy, the traditional Turkana diet is extremely protein-rich: 70-80% of calories are derived from milk or other animal products [51]. For detailed descriptions of the diet, climate, and lifestyle experienced by traditional, pastoralist Turkana, see work from the South Turkana Ecosystem Project [52].
Over the last several decades, many urban areas in central Kenya have experienced an influx of Turkana people as a result of country-wide infrastructure improvements and rapid cultural, economic, and social changes; for the same reasons, the capital of Turkana county (Lodwar) has also become increasingly urban and market-integrated. In our study, we use the term “urban” to refer to people living in densely populated cities characterized by many permanent businesses, and where most people engage with the market economy and/or work wage labor jobs. We defined “urban” individuals as those that no longer practice pastoralism and reside in one of three cites included in our study—Nanyuki, Lodwar, and Kitale. All three of these cities have population sizes >20k and are among the top 50 largest cities in Kenya (https://worldpopulationreview.com/). We also included non-pastoralists residing in suburbs in Laikipia county in central Kenya in the urban category, because Laikipia is a cosmopolitan area with several large cities (e.g., Nanyuki, Nyahururu, and Rumuruti).
We defined pastoralists as residents of Turkana county who self-reported their main subsistence activity as “pastoralism”, who owned livestock, and who drink milk every day (i.e., they rely on their livestock for subsistence). In previous work, we also defined a third category of Turkana who no longer practice pastoralism but still live in the relatively remote and rural Turkana homelands [36]. For the purposes of this study, we focused only on the extremes of the Turkana lifestyle spectrum (pastoralist versus urban) because SES is more difficult to define and operationalize in the intermediate context, where both livestock and material goods contribute to wealth (see next section).
Data were collected in Turkana, Laikipia, and Trans-Nzoia counties between April 2018 and February 2020. During this time, researchers visited locations where Turkana individuals were known to reside (Figure S1). At each sampling location, local chiefs and elders were first consulted about the project. If they believed the study to be of interest to their community, a larger meeting was held to explain the project to all interested individuals. After this period of discussion, adults (>18 years old) of self-reported Turkana ancestry were invited to participate in the study. The study involved a structured interview, blood sample collection, and anthropometric measurements. Additional background on the study as well as detailed sampling procedures and demographic summaries are provided in [36].
Structured interviews
Self-reported health, potential behavioral mediators, and covariates
Structured interviews were conducted with all participants to collect information about demography, reproductive history, diet, early life experiences, lifestyle, and wealth. All interviews were conducted in a language familiar to the participant (English, Turkana, or Swahili). The following self-reported variables from the interviews are relevant to our analyses:
Sex
Age
Main subsistence activity, chosen from the following categories: self-employment, formal employment, petty trade, farming, pastoralism, hunting and gathering, other
Highest education level scored as follows: 0=none, 1=lower primary school, 2=upper primary school, 3=secondary school, 4=education beyond secondary school
Number of surviving children
Number of wives (for men only)
Whether the participant used medications or sought medical treatment when ill in the last month (Y/N/NA, if not ill in the last month)
Whether the participant used alcohol, tobacco, or cigarettes (never/occasionally/daily)
Whether the participant was currently fasting (this covariate was included in analyses of blood glucose)
Whether the participant experienced each of the following health issues in the last 3 months (Y/N for each question): swollen extremities, fatigue or weakness, shortness of breath, diarrhea, worm infection, stomach pain, vomiting, constipation, coughing, difficulty breathing, dizziness, headaches, chest pain, bruises, cuts and scrapes, or burning during urination.
We also used a food frequency questionnaire to collect information about the consumption of meat, milk, bread, sugar, salt, and cooking oil. We focused on these items because they reflect foods that are essential (meat, milk) or uncommon (bread, sugar, salt, cooking oil) in the diet of traditional pastoralists. Participants were asked how often a specific item was used or consumed and were given the following answer choices: never, rarely, 1-2 times per week, >2 times per week, or every day. These answers were converted to a scale of 0-4. Results for sugar, salt, and cooking oil were tallied and combined into a single metric because these answers were highly correlated among both pastoralists and urban individuals (all R2>0.9).
Because 1) we were interested in a life course perspective on social conditions and health and 2) early life challenges can set individuals on a course toward adverse sociobehavioral environments in adulthood [43–45], we interviewed participants about their early life experiences, adapting the CDC’s Adverse Childhood Experiences (ACEs) instrument [53]. Specifically, we created a tally of the number of adversities each participant experienced as a child (<12 years old) from the following list: mother or father absent (i.e., didn’t leave with the participant when they were young due to death, abandonment, or other circumstances), verbal abuse or threat of violence by a caregiver, physical abuse by a caregiver, witness of verbal or physical abuse toward mother, exposure to mental illness, exposure to alcoholism or other substance abuse, and food insecurity. These questions were asked using the phrasing provided by the Centers for Disease Control (https://www.cdc.gov/violenceprevention/aces/ace-brfss.html) (modeled after [53]), except for the food insecurity question, which we added because it is a common concern in Kenya. We also asked participants where they were born, as well as what the main subsistence strategy and occupation of their parents was during childhood.
SES/material wealth metrics
We drew on our interview data to create two separate metrics of SES, one for the pastoralist context and for the urban context. Our measures are meant to capture key indicators of absolute material wealth in each context, rather than other potential dimensions of social status such as influence or standing in the community (as perceived by the focal individual or other community members), embodied wealth, or inequality/relative material wealth. Defining absolute material wealth in itself is a complex and challenging task, and many approaches have been taken [54,55]. Our approach is based on the data that were available as well as precedent in the literature.
Livestock are the primary source of material wealth among pastoralists [27]. Therefore, in the pastoralist setting, we defined SES as the total number of multispecies livestock owned by the household the individual belonged to, following [27] and references therein. This value was log2 transformed because of skew. Importantly, this measure of SES was strongly correlated with other possible metrics of material wealth [32,56], for example livestock translated into estimated market prices (β=1.01, p-value<10−16, linear regression) as well as the ratio of total multispecies livestock holdings to the number of household members (β=0.913, p-value<10−16, linear regression).
In the urban setting, we used a tally of durables/goods, dwelling characteristics, and other household assets as an index of SES and absolute material wealth. This approach is relatively common [54] and the specific list was derived from previous work [57–59]. We tallied household possession of the following items to create an index ranging from 0-11: finished floor, finished roof, electricity, television set, mobile phone, flush toilet, gas cooking, indoor tap water, treated water, >1 room in the household, and <= 2 household members per room. We did not include durables and dwelling characteristics in the pastoralist SES index, because very few individuals in the pastoralist setting own any of goods listed above (3.1%). Similarly, we did not include livestock holdings in our urban SES metric, because few individuals in this setting own livestock (21.4%). For the subset of urban individuals that own livestock, livestock holdings are strongly correlated with the urban SES index, suggesting we are not missing primary sources of wealth by not counting livestock (β=0.027, p-value=0.036, Poisson regression). Our urban SES index is also strongly associated with education levels (β=0.203, p-value<10−16, Poisson regression), another common measure of SES in HICs and industrialized settings [60].
Measuring biomarkers of cardiometabolic health
In addition to collecting data on self-reported health, we also measured 10 biomarkers of cardiometabolic health. We used standard anthropometric approaches to measure body mass index (BMI), waist circumference, body fat percentage, as well as systolic and diastolic blood pressure. We also collected venous blood to measure blood glucose levels, total cholesterol, triglycerides, and high- and low-density lipoproteins (HDL, LDL). The collection of all of these measures are described in detail in [36].
Data were excluded in a handful of cases where individuals were extreme outliers, indicating a likely error (>5 standard deviations from the mean). Prior to statistical analyses, all biomarkers and SES measures (when they were used as predictor variables) were mean-centered and scaled by their standard deviation using the “scale” function in R [61]. Consequently, all effect sizes reported in the methods are standardized, and represent the effect of a given variable on the outcome in terms of increases in standard deviations.
Measuring serum cortisol
Venous blood was collected from each participant in a serum separator tube (Fisher Scientific) and spun immediately for 15 minutes at 2500 RPM in a portable centrifuge. The serum layer was then pulled off the top of the tube, transferred to a 2mL cryovial, and frozen at - 10C in a portable freezer. Samples were kept in the portable freezer for no longer than one week, after which they were transferred to long-term storage in a -20C freezer at Mpala Research Centre (Laikipia, Kenya). Samples were exported to the United States on dry ice, and upon arrival were stored at -80C.
216 serum samples were thawed on ice and used to quantify cortisol with the Cortisol Elisa Assay Kit from Eagle BioSciences, according to the manufacturer’s instructions. Samples were randomized across three plates, and the R2 between the expected and observed concentrations for the calibrator curves were 0.9994, 0.9989, and 0.9977. Two control samples of known concentration were run in duplicate on each plate, and all values fell within the acceptable range specified by the kit’s manufacturer. In the total sample, we did not find any evidence for plate effects (ANOVA, F=1.328, p-value=0.267, n=216) and we did observe expected effects of age, sex, and time of day of sample collection [62–64]. Specifically, our linear models revealed that males exhibited higher cortisol levels than females (β=-0.484, p-value=0.013), cortisol levels increased with age (β=0.021, p-value=0.006), and cortisol levels were higher in the morning (β=-0.104, p-value=0.018).
Testing for SES effects on health in the pastoralist and urban settings, and exploring potential mediators
For each of the 16 self-reported health measures, we used binomial regression to ask whether SES was predictive of health in each context. For each of the 10 biomarkers of cardiometabolic health, we used linear models to ask whether SES was predictive of health outcomes in the urban or pastoralist setting (analyzed separately; see Table S1 for sample sizes). We did not run models in cases where <1% of people experienced a given self-reported health issue (Table S1). All analyses controlled for self-reported age and sex as fixed effects covariates. For each health outcome, we also ran analyses that considered a sex x SES interaction term, and we used the results of the second model if the delta AIC between this model and the first model was >2 (Table S1). All p-values were corrected for multiple hypothesis testing using a Benjamini-Hochberg false discovery rate approach [65]. We considered SES effects to be significant in a given context at a 10% FDR cutoff.
We also explored potential mediators of SES-health associations. To do so, we used formal mediation analyses [40,41] following the methods in [5,36,66]. We considered the following behavioral factors as potential mediators: cigarette smoking (Y/N), alcohol usage (Y/N), tobacco usage (Y/N), use of health care resources (i.e., whether the participant used medications or sought medical treatment when ill; Y/N/NA), frequency of use of salt/sugar/oil in cooking (0-4), and frequency of consumption of bread, meat, or milk (0-4). We also considered cortisol levels as a potential biological mediator. For a variable to be a potential mediator, it needs to be correlated with the predictor variable of interest. We therefore used linear, binomial, and Poisson regression (for continuous, binary, and count variables, respectively) to predict each mediator as a function of SES, in the pastoralist and urban samples, respectively. All models controlled for age and sex, and the cortisol models also controlled for time of day of sample collection.
We tested all variables for mediation that were predicted by SES at a relaxed nominal p-value of 0.1, and that were associated with SES in a direction that made sense for mediating health effects. To do so, we fit two models: 1) an “unadjusted” model that included the effect of the predictor variable on the outcome (i.e., the effect of SES on a given health outcome, controlling for covariates) and 2) an “adjusted” model that is identical to model 1 but also includes the putative mediator as a covariate. If the predictor’s effect on the outcome is explained by the mediator, then the effect of the predictor (β SES) will decrease when the mediator is included in the adjusted model and absorbs variance otherwise attributed to the predictor. To assess significance, we estimated the decrease in β SES between the unadjusted and adjusted models across 1000 iterations of bootstrap resampling. We considered a variable to be a significant mediator if the lower bound of the 95% confidence interval for the decrease in β SES did not overlap with 0.
Testing for SES effects on reproductive success in the pastoralist and urban settings
We were also interested in the potential fertility consequences of SES, as identifying these links is important for thinking about how SES effects on health may ultimately impact Darwinian fitness, and thus for informing our understanding about how SES-associated traits (e.g., striving for wealth or other forms of status) evolve in humans [39]. Therefore, we used Poisson regression to predict the number of surviving offspring each individual had as a function of SES, age, sex, and the interaction between SES and sex (as this improved model fit in both contexts). The total number of surviving offspring was self-reported, and was not available in terms of offspring sex or age breakdowns (e.g., we could not calculate number of offspring surviving past a certain age). We also note that number of surviving offspring is not the same as Darwinian fitness, which is difficult to measure, but it is a commonly used proxy in the literature and is routinely considered a fitness-related trait (e.g., [39]).
For men in the pastoralist setting, we also ran a supplementary analysis to understand whether SES effects on reproductive success were mediated by SES effects on number of wives. In traditional Turkana culture, livestock are used as bridewealth and polygyny is common [29]. This practice is much less common in urban settings: 42% versus 9% of married men reported >1 wife in the pastoralist and urban settings, respectively. To test whether SES effects on reproductive success were mediated by SES effects on number of wives, we fit two Poisson regression models to data from male pastoralists: 1) an “unadjusted” model that included the effect of SES on number of children, controlling for age and 2) an “adjusted” model that was identical to model 1 but also included number of wives as a covariate. We then used the same bootstrap approach described above to assess significance.
Understanding the relationships between lifestyle, early life adversity, adult SES, and adult health
To test whether early life adversity (ELA) predicted adult SES in each context, we used linear regression (for pastoralists) and Poisson regression (for urban individuals) to predict each wealth index as a function of our cumulative ELA score controlling for age and sex. Cumulative ELA was considered on a scale of 0-5, with individuals experiencing >5 adversities collapsed into the 5 category to prevent the influence of outliers. We also used Poisson regression to understand whether cumulative ELA itself was predicted by 1) age, sex, or lifestyle (urban versus pastoralist) in the total sample and 2) age, sex, or parental subsistence strategy (coded as pastoralist, other, or formal employment) in the pastoralist sample only.
To test whether ELA was related to adult health in each context, we used linear regression controlling for age and sex to predict our 10 biomarkers of cardiometabolic health as a function of cumulative ELA. Unfortunately, we did not conduct interviews on self-reported health and early life experiences for the same set of individuals, and we therefore could not assess the relationships between these measures. All p-values were corrected for multiple hypothesis testing and were considered significant if they passed a 10% FDR.
Ethics approval
This study was approved by Princeton University’s Institutional Review Board for Human Subjects Research (IRB# 10237), and Maseno University’s Ethics Review Committee (MSU/DRPI/MUERC/00519/18). We also received county-level approval for research activities, and research permits from Kenya’s National Commission for Science, Technology and Innovation (NACOSTI/P/18/46195/24671). Written, informed consent was obtained from all participants after the study goals, sampling procedures, and potential risks were discussed with community elders and explained to participants in their preferred language (by both a local official, usually the village chief, and by researchers or field assistants).
Results
Social status effects on health and a fitness-related trait are highly context-dependent
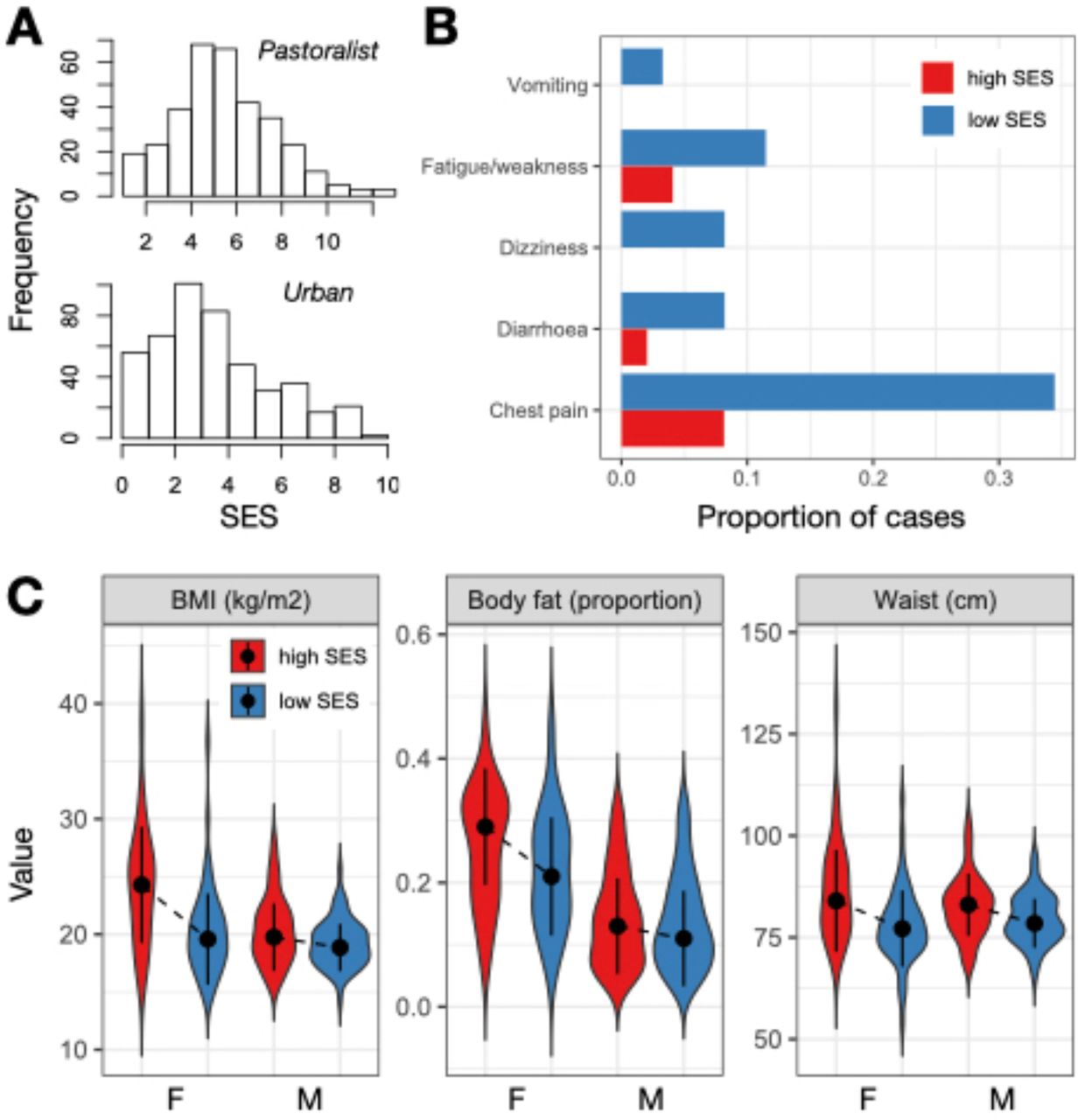
Among traditional pastoralists, SES was positively associated with self-reported health. High status individuals were significantly less likely to suffer from recent chest pain (β SES=-0.624, q-value=0.016), diarrhea (β SES=-1.472, q-value=0.057), vomiting (β SES=-2.604, q-value=0.074), dizziness (β SES=-1.424, q-value=0.016), and fatigue or weakness (β SES=-1.094, q-value=0.016; Table S1-3). These effect sizes were substantial: for example, 33%, 14%, and 9% of individuals in the lowest quartile of the SES distribution experienced chest pain, fatigue/weakness, and dizziness in the past three months, compared to 8%, 7%, and 0% of individuals in the highest quartile (Figure 1B). There were no significant relationships between SES and measures of cardiometabolic health among traditional pastoralists (Table S1); most pastoralists were cardiometabolically healthy (Figure S2).
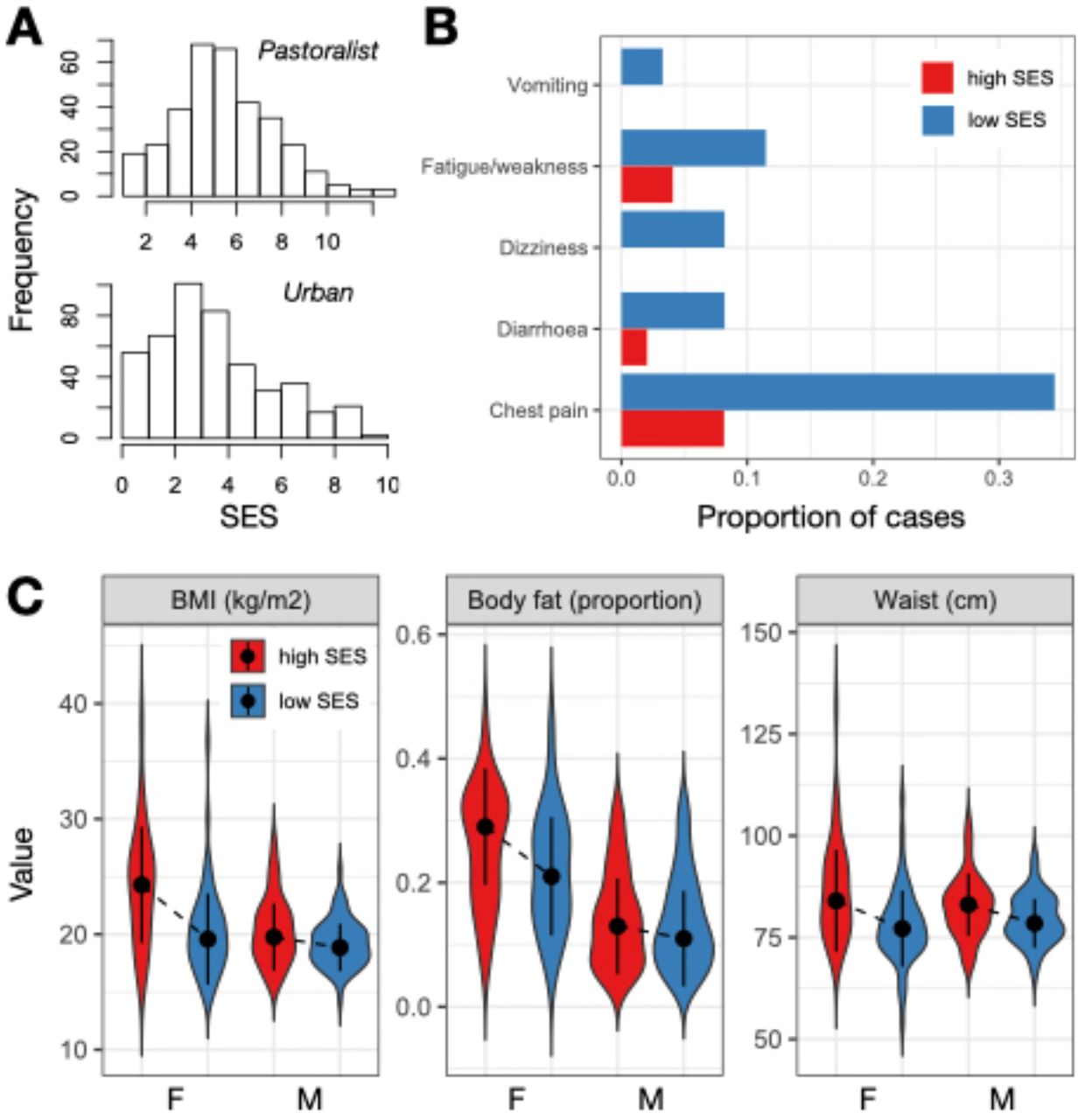
(A) Distribution of SES measures among pastoralists (where SES was defined as log2 transformed livestock counts) and urban individuals (where SES was defined by a tally of market-derived possessions). (B) Proportion of people reporting various health issues as a function of SES in the pastoralist setting. For visualization, data from individuals in the highest and lowest SES quartiles are plotted. (C) Distribution of cardiometabolic biomarker values as a function of sex and SES (highest versus lowest quartiles) in the urban setting. Dots represent the median of each distribution and solid lines represent the median +/- 1 standard deviation.
The direction of the effect of SES on health was reversed in the urban setting and captured entirely different health outcomes. In this context, we found no relationships between SES and self-reported health, even though the frequency of self-reported health issues was higher than in the pastoralist setting (Table S1). Among urban individuals, high status was associated with biomarkers of worse cardiometabolic health: higher BMI (β SES=0.425, q-value=1.02×10− 7), waist circumference (β SES=0.436, q-value=7.46×10−9), diastolic blood pressure (β SES=0.113, q-value=0.088), and body fat (β SES=0.215, q-value=5.82×10−4; Table S1-3). With respect to BMI, a modest number of individuals in the urban setting met the criteria for being overweight (12.56%) or obese (2.90%); nevertheless, BMI is often a linear predictor of type II diabetes [67] and cardiovascular disease [68] (though not necessarily of mortality [69]), suggesting that this SES-driven variation is still potentially meaningful. For all of the above traits except blood pressure, we observed that women were more sensitive to SES effects on health (body fat: β SES x sex=-0.157, p-value=0.03; waist circumference: β SES x sex=-0.225, p-value=0.012; BMI: β SES x sex=-0.313, p-value=8.63×10−4; Figure 1C).
In both the pastoralist and urban settings, SES was associated with reproductive success, but in opposite directions. In the urban setting, high SES was associated with fewer surviving offspring (β SES= -0.129, p-value=8.68×10−4), and an interaction effect pointed to this effect being primarily driven by stronger effects in women than men (β SES x sex=0.136, p-value=0.010; Figure 2 and Table S4). In the pastoralist setting, a main effect of SES was not significant (β SES=-6.86×10−3, p-value=0.846), but an interaction term supporting stronger SES effects in men than women was significant (β SES x sex=0.095, p-value=0.047; Figure 2 and Table S4). Causal mediation analyses revealed that the effect of male pastoralist SES on reproductive success is partially explained by SES effects on the number of wives a man had (estimated proportion of the total effect that is mediated=25%, p-value=0.046; Figure S3).

(A-B) Number of living children as a function of age among pastoralist women and men in the highest versus lowest SES quartiles. (C-D) Number of living children as a function of age among urban women and men in the highest versus lowest SES quartiles.
Mediators of the SES-health relationship among pastoralist and urban Turkana
We explored the role of cortisol as well as sociobehavioral variables in mediating links between SES and health. We did not find any association between SES and serum cortisol in either the pastoralist (β SES=0.103, p-value=0.577, N=79) or urban samples (β SES=0.025, p-value=0.887, N=59) (Figure S4). Therefore, we did not test this variable for mediation. In terms of sociobehavioral variables, several of the factors we hypothesized might explain social gradients in health were also not predicted by SES, and were therefore excluded from further mediation analyses. In particular, SES was unrelated to the usage of cigarettes, alcohol, tobacco, health care resources, meat, and bread in the pastoralist setting (Table S5). In the urban setting, SES was unrelated to usage of cigarettes and health care resources (Table S5-6). SES was associated with alcohol (β SES=-0.254, p-value=0.019) and tobacco usage (β SES=-0.658, p-value=3.56×10−4) among urban Turkana, but the direction of these effects was not compatible with mediation (i.e., low SES individuals exhibited worse health habits).
Because they were significantly or marginally associated with SES, we tested whether the following variables could explain observed SES-health associations: 1) greater usage of salt, sugar, and oil among low SES pastoralists (β SES=-1.304, p-value<2×10−16) and 2) greater consumption of meat (β SES=0.150, p-value=1.29×10−5), bread (β SES=0.150, p-value=2.18×10− 13), and milk (β SES=0.141, p-value=3.13×10−5), as well as greater reliance on salt, sugar, and oil (β SES=0.026, p-value=0.097), among high SES individuals in the urban setting (Table S5). We found generally minimal evidence for mediation (Table S7), with the exception that salt, sugar, and oil consumption explained an estimated 11% of the effect of SES on waist circumference in urban individuals.
Lifestyle patterns early life experiences, but early life adversity does not predict adult SES or adult health
Turkana practicing pastoralism as adults experienced greater cumulative early life (ELA) relative to those living in urban settings in adulthood (β lifestyle=-0.465, p-value=1.97×10−3; Table S8). We also found that a lifestyle x sex interaction improved model fit, with the direction of this effect suggesting that pastoralist men experienced the highest levels of cumulative ELA (β lifestyle x sex=-0.343, p-value=0.075; Figure 3). For example, 11.1% and 15.6% of pastoralist women and men experienced 5 or more adversities, while 6.3% and 2.9% of urban women and men experienced the same level of hardship. For reference, these numbers are estimated at 10.3% and 6.9% for women and men in the United States [70] (though we note the US numbers are derived from a slightly different questionnaire; Table S9 and Figure S5). Age of the study participant also trended toward a positive association with cumulative ELA in both contexts, suggesting that incidence of ELA has generally reduced over time (β age=0.007, p-value=0.058).
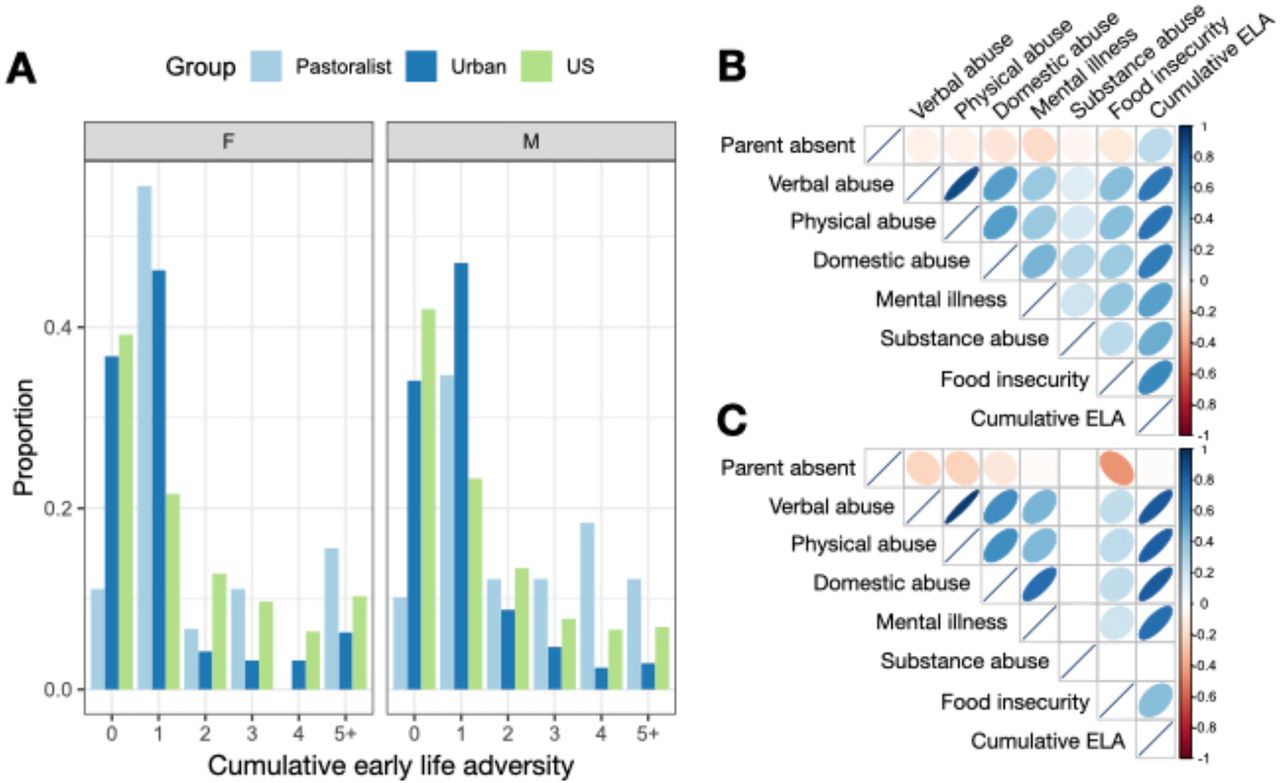
(A) Number of early life adversities experienced by sex and population. US data were sourced from [70]. Correlations among individual sources of early life adversity as well as cumulative early life adversity tallies for (B) urban and (C) pastoralist individuals. Note that no pastoralist individuals were exposed to substance abuse within their household growing up, and no pairwise correlations are therefore presented that involve this measure.
While individuals practicing pastoralism as adults all grew up in pastoralist families, 46% of urban adults grew up in pastoralist families; the remaining 30% and 24% had parents that relied on formal employment or other types of employment, respectively. We found that, within urban adults specifically, early life subsistence strategy mattered for ELA in ways that were consistent with results from the total sample: individuals who grew up in pastoralist families experienced more ELA than individuals whose parents were formally employed (β parental occupation=-0.275, p-value=0.036). We did not find any significant differences for the other contrasts (i.e., formal employment versus other employment or pastoralism versus other employment, both p>0.05).
While cumulative ELA is clearly patterned by lifestyle, we did not find any compelling evidence that this variation matters for adult health in our sample (Table S10). Controlling for age and sex, we also did not find that cumulative ELA was predictive of adult SES in the urban setting (β ELA=0.031, p-value=0.219). In the pastoralist setting, we observed a trend toward greater cumulative ELA predicting higher adult social status (β ELA=0.268, p-value=0.061). The direction of this marginal effect was unexpected and suggests that cumulative ELA does not mediate the (positive) effects of SES on health we observe in traditional pastoralists.
Discussion
Here, we test the hypothesis that shifts toward more urban lifestyles alter or exacerbate the relationship between SES and health. Previous work has suggested that the effects of social status on health may be exacerbated in HICs relative to ancestral and pre-industrial human societies [13]. In particular, the social conditions under which humans evolved were thought to feature less rigid hierarchies, greater kin support, and greater upward mobility than what is currently observed in HICs [13,15,16]. They also did not routinely feature socially stratified health care resources, structural racism, and other forms of institutionalized inequality [17–20]. Thus, the negative effects of low social status and limited material wealth that are common in HICs could be a recent byproduct of post-industrial societal, economic, or even epidemiological changes (e.g., most deaths in HICs are driven by non-communicable rather than infectious diseases, which could have different relationships to SES). Identifying the degree to which this hypothesis is true is important for understanding the evolution of social gradients in health as well as the best strategies for combating them.
We found strong evidence that the recent transition from pastoralism to an urban, market-based lifestyle alters the relationship between social status and health among the Turkana. Among traditional Turkana pastoralists, high SES is associated with better self-reported health, but unassociated with cardiometabolic health. In contrast, among urban Turkana, high SES is associated with worse cardiometabolic health. At first glance, these results do not necessarily adhere to the framework laid above, which would predict that low SES would be associated with worse health in both the pastoralist and urban setting, with these effects magnified in the urban setting. Instead, we find that the presence and direction of SES effects are modified as a function of lifestyle. We also found similar opposing effects when examining a fitness-related trait: high SES predicts more offspring in the pastoralist setting, partially driven by wealth effects on polygyny, but fewer offspring in the urban setting, where polygyny is uncommon and greater wealth is likely associated with later marriage and family planning [71].
While some of our results run counter to the large literature linking low SES to worse outcomes in HICs, they highlight the importance of socioecological context when interpreting social gradients in health. In the pastoralist setting, wealth is likely channeled into traditional foods, family growth, and other resources with positive effects on self-reported health, while in the urban setting, wealth is likely channeled into consumption, purchase, and amenities that negatively impact cardiometabolic health (e.g., processed foods, vehicles to minimize physical activity). We found some support for this idea in our mediation analyses, where increased consumption of market-derived foods—namely, salt, sugar, and oil—mediated 11% of the effect of SES on waist circumference in the urban setting. We note that the idea that high socioeconomic status in newly urban Turkana translates into purchases that negatively impact cardiometabolic health (e.g., processed foods) is also consistent with literature on epidemiological and nutritional transitions, in which newly developed countries often suffer an increased burden of non-communicable diseases [72].
The highly context-dependent nature of SES effects observed in the Turkana dovetails with recent work in non-human primates, which has also found that both the magnitude and direction of social status effects varies across systems. For example, work in humans and captive female macaques has emphasized the association between low social status and increased expression of innate immune and inflammation-related genes [5,73,74]. However, in wild baboons, weak effects in the expected direction were observed in females [12], while strong patterns in the opposite direction were observed in males [66] (i.e., high social status predicted increased expression of inflammation-related genes). Importantly, social status is attained through direct physical competition in male baboons but not in any of the other species or sexes, suggesting that the heterogeneity between studies reflects differences in the nature of social hierarchies. Such a nuanced picture is consistent with decades of work on non-human primate social systems that has emphasized the diverse ways in which status is attained and maintained across systems, as well as the diverse sets of costs and benefits that are associated with status [7,16]. In these systems, it appears that the physiological and health correlates of social status vary as a function of the specifics of the social environment as well. Our results suggest that the same logic could be applied toward thinking about heterogeneity in SES gradients in health in human societies.
In addition to showing that transitions to urban, market-integrated lifestyles alter the relationship between wealth and health in a single population, our study also reports novel findings related to early life experiences. In particular, we used the adverse childhood experiences (ACEs) framework to document variation in early life adversity (ELA) as a function of lifestyle. We found greater ELA in the traditional, pastoralist setting, but no relationship between ELA and adult SES or health in either setting. The lack of an association with health is noteworthy given the large body of literature linking greater ACE exposure to earlier death and later life cardiovascular, autoimmune, and neurodegenerative diseases in HICs [53,75–82].
However, there has been very little work on the subject in low and middle income countries (but see [83,84]), despite childhood adversity being a growing area of international interest [85]. It may be that larger sample sizes are needed to uncover effects in the Turkana, that early life adversity does not carry the same psychosocial and physiological costs as in HICs because of differing cultural norms [86], or that the ACE questionnaire does not capture the most salient types of adversity experienced by the Turkana (e.g., experiences of livestock loss or raiding in the traditional context [35,86,87]). In general, more research is needed that considers a wider variety of social, economic, and cultural circumstances in the study of early life adversity.
There are several limitations to the present study, as well as open directions for future work. First, our analyses of SES effects on serum cortisol levels did not reveal any significant associations and were limited by a small sample size. Previous work in human and non-human primates has shown that low social status individuals are often chronically stressed, which leads to altered hypothalamic-pituitary-adrenal (HPA) axis function reflected in chronically elevated cortisol levels [6,7,16]. However, social status effects on cortisol are also known to be context-dependent across non-human primate hierarchies [88], suggesting that this may be another area where lifestyle modifies the direction or magnitude of SES effects. Future work could expand the sample set and explore the context-dependency of cortisol levels across lifestyle groups. Another limitation of the current study is that we were unable to identify sociobehavioral mediators of the SES-health relationship in either the pastoralist or urban setting. It may be that these relationships are not mediated by indirect effects of behavior, and are instead entirely explained by direct effects of SES on biological mechanisms we have yet to uncover (e.g., cortisol levels) or explore (e.g., gene regulation [5,66,73,89]). Alternatively, SES effects may be mediated by behavioral variables that were not captured by our surveys. For example, the relationship between social status, stress, and health may critically depend on the availability of kin and social support [6,28,90], which we did not measure here and which likely varies dramatically as a function of lifestyle.
Data Availability
anonymized data underlying the main results will be publicly available on Dryad following publication.
Data Availability
anonymized data underlying the main results will be publicly available on Dryad following publication.
Funding and conflicts of interest
This work was supported by awards to J.F.A. through Princeton University’s Dean for Research Innovations Funds as well as the Chan Zuckerburg Initiative. A.J.L. was supported by a postdoctoral fellowship from the Helen Hay Whitney Foundation. We declare no conflicts of interest.
Acknowledgments
We thank previous members of the Turkana Health and Genomics Project for their contributions, especially S. Lowasa, D. Mukhongo, S. Ngatia, E. Loowoth, and M. Ndegwa. We thank J. Orina, K. Tombak, and D. Rubenstein for logistical help in Kenya and members of the Graham lab at Princeton for technical assistance with the cortisol Elisa assay. We are also grateful to the staff of Mpala Research Centre for their essential support, especially F. Hassan, C. Nzomo, B. Wanjohi, G. Chege, T. Maina, and J. Nakolonyo. Finally, we thank all of the Turkana communities for their contributions to and support of our scientific work.
References
- 1.↵
- 2.
- 3.↵
- 4.↵
- 5.↵
- 6.↵
- 7.↵
- 8.↵
- 9.↵
- 10.↵
- 11.
- 12.↵
- 13.↵
- 14.↵
- 15.↵
- 16.↵
- 17.↵
- 18.
- 19.
- 20.↵
- 21.↵
- 22.
- 23.
- 24.
- 25.
- 26.↵
- 27.↵
- 28.↵
- 29.↵
- 30.↵
- 31.↵
- 32.↵
- 33.↵
- 34.↵
- 35.↵
- 36.↵
- 37.↵
- 38.↵
- 39.↵
- 40.↵
- 41.↵
- 42.↵
- 43.↵
- 44.
- 45.↵
- 46.↵
- 47.
- 48.↵
- 49.↵
- 50.↵
- 51.↵
- 52.↵
- 53.↵
- 54.↵
- 55.↵
- 56.↵
- 57.↵
- 58.
- 59.↵
- 60.↵
- 61.↵
- 62.↵
- 63.
- 64.↵
- 65.↵
- 66.↵
- 67.↵
- 68.↵
- 69.↵
- 70.↵
- 71.↵
- 72.↵
- 73.↵
- 74.↵
- 75.↵
- 76.
- 77.
- 78.
- 79.
- 80.
- 81.
- 82.↵
- 83.↵
- 84.↵
- 85.↵
- 86.↵
- 87.↵
- 88.↵
- 89.↵
- 90.↵





{kind=link}
{kind=link}
{kind=link}