
Abstract
One of the defining criteria of Variants of Concern (VOC) is their ability to evade pre-existing immunity, increased transmissibility, morbidity and/or mortality. Here we examine the capacity of convalescent serum, from a well defined cohort of healthcare workers (HCW) and Patients infected during the first wave from a national critical care centre in the UK, to neutralise B.1.1.298 variant and three VOCs; B.1.1.7, B.1.351 and P.1. Furthermore, to enable lab to lab, country to country comparisons we utilised the World Health Organisation (WHO) International Standard for anti-SARS-CoV-2 Immunoglobulin to report neutralisation findings in International Units. These findings demonstrate a significant reduction in the ability of first wave convalescent plasma to neutralise the VOCs. In addition, Patients and HCWs with more severe COVID-19 were found to have higher antibody titres and to neutralise the VOCs more effectively than individuals with milder symptoms. Widespread use of the WHO International Standard by laboratories in different countries will allow for cross-laboratory comparisons, to benchmark and to establish thresholds of protection against SARS-CoV-2 and levels of immunity in different settings and countries.
Introduction
SARS-CoV-2 is the causative agent of the COVID-19 global pandemic, resulting in more than 125 million cases and over 2.5 million deaths (1). Since the start of the outbreak in late 2019, the extensive sequencing of circulating virus has revealed the gradual evolution of variants, emerging independently in many countries around the world. Coronaviruses are enveloped viruses with single stranded positive sense RNA genomes ranging from 26 to 32 kilobases in length. SARS-CoV-2 is a member of the β-coronavirus genus which also comprises SARS-CoV (2) and Middle East respiratory syndrome coronavirus (MERS-CoV) (3). As the pandemic progressed a number of single amino acid mutations in the Spike protein were detected, such as D614G and A222V. The D614G mutation was found to increase the density of Spike protein on virions and increased infectivity (4). The rise of variants in circulation containing several mutations in the viral genome alter several properties of the virus (5). Due to a number of criteria including, increased transmissibility, mortality or morbidity, and the ability to evade natural immunity, these variants have been designated as Variants of Concern (VOC). Some of these mutations found in the N-terminus and receptor-binding domain (RBD) of the Spike protein are associated with immune evasion (6–8). For instance, the E484K mutation in the RBD of the B.1.351 and P.1 VOCs has been reported to cause up to a ten-fold reduction of neutralisation (9), while in the more recent B.1.427/B.1.429 detected in California, USA, the L452R mutation resulted in a 4 fold reduction (10). Antibodies generated from prior infection or vaccination against the initial virus isolates may provide reduced protection, giving rise to subsequent waves of infection in regional populations previously impacted by earlier COVID-19 outbreaks (11,11–13).
The first notable SARS-CoV-2 variant was linked to an outbreak on a mink farm in Denmark, resulting in a culling program to mitigate risk of spreading (14,15). Referred to as cluster 5 or B.1.1.298, several different clusters of mutations were identified, with the most abundant population containing missense and deletion mutations on the Spike; 69/70del, Y453F and D614G. Shortly after, in September 2020, a new variant was detected in the United Kingdom designated B.1.1.7 which was reported to be more transmissible (16,17). In December 2020, the rise of a new variant designated as B.1.351 was detected in South Africa. This new variant has the E484K mutation in the Spike protein that is believed to have a strong impact on antibody evasion (9). Lastly, in Manaus, Brazil, a variant designated P.1 was detected, which also harboured mutations similar to B.1.351, and has been reported to also evade antibodies in previously infected individuals (11,13,18). It is of high importance to assess the effectiveness of antibodies from individuals who have recovered from natural infection, as this would allow us to ascertain whether natural infection from the early Wuhan virus isolates, herein referred to as B.1, may offer protection against the newly circulating VOCs, as well as assessing the efficacy of neutralising antibodies generated from vaccines. Having this information would be very informative to develop our understanding of SARS-CoV-2 immune correlates of protection (Figure 1), since neutralising antibody levels are predictive of immune protection (19,20).

To prevent laboratory to laboratory variability between assays, the International Standard was created to standardise results which would allow for cross laboratory comparisons. With gradual accumulation of data, this would permit further analysis into determining correlates for protection against SARS-CoV-2.
Here, we assessed antibodies from plasma from convalescent Health Care Workers (HCWs) and patients who were infected during the first wave in the United Kingdom in early 2020 (21). Using well defined and cross validated lentiviral based pseudotyped viruses bearing the Spike protein from SARS-CoV-2 B.1 and the three VOCs, neutralisation assays were performed, reporting IC50 values in International Units (IU) according to WHO recommendations (22).
Materials and Methods
Tissue Culture
Human Embryonic Kidney 293T/17 (HEK293T17) cells were maintained in DMEM with 10% foetal bovine serum, 1% penicillin/streptomycin and incubated at 37°C and 5% CO2.
Plasma Collection
Serum and plasma samples were obtained from HCWs and patients referred to the Royal Papworth Hospital, Cambridge, UK (RPH) for critical care. COVID-19 patients hospitalised during the first wave and as well as NHS healthcare workers working at RPH served as the exposed HCW cohort (Study approved by Research Ethics Committee Wales, IRAS: 96194 12/WA/0148. Amendment 5). NHS HCW participants from the Royal Papworth Hospital were recruited through staff email over the course of 2 months (20th April 2020-10th June 2020) as part of a prospective study to establish seroprevalence and immune correlates of protective immunity to SARS-CoV-2. Patients were recruited in convalescence either pre-discharge or at the first post-discharge clinical review. All participants provided written, informed consent prior to enrolment in the study. Plasma from NHS HCW and patients used in this study were collected between July and September 2020, approximately 3 months after they were enrolled in the study. Clinical assessment and WHO criteria scoring of severity for both patients and NHS HCW was conducted following the ‘COVID-19 Clinical Management: living guidance (https://www.who.int/publications/i/item/WHO-2019-nCoV-clinical-2021-1). Scoring is based on progression of respiratory disease and cardiovascular collapse: 1=asymptomatic; 2=mild disease; 3=moderate pneumonia; 4 = severe pneumonia; 5=adult respiratory distress syndrome; 6=sepsis; 7= septic shock.
For cross-sectional comparison, representative convalescent serum and plasma samples from seronegative HCWs, seropositive HCW and convalescent PCR-positive COVID-19 patients. The serological screening to classify convalescent HCW as positive or negative was done according to the results provided by a CE-validated Luminex assay detecting N-, RBD- and S-specific IgG, (23) a lateral flow diagnostic test (IgG/IgM) and an Electro-chemiluminescence assay (ECLIA) detecting N- and S-specific IgG. Any sample that produced a positive result by any of these assays was classified as positive. The clinical signs of the individuals from which the sample was obtained ranged from 0 to 7 according using the WHO classification described above. Thus, the panel of convalescent serum samples (3 months post-infection) were grouped in three categories: a) Patients (n=38); b) Seropositive Staff (n=24 samples); and c) Seronegative Staff (n=39). Age, Sex and Symptom severity score is shown in Table 1.
Cohort Demographic and Severity Score Classification.
*Symptom severity score: ‘COVID-19 Clinical Management: living guidance (https://www.who.int/publications/i/item/WHO-2019-nCoV-clinical-2021-1).
Generation of Spike expression plasmids
The B.1 SARS-CoV-2 Spike expression plasmid (pcDNA3.1+) is based on the Wuhan-Hu-1 sequence and was kindly gifted by Professor Xiao-Ning Xu, Imperial College, London. Mutations of each variant sequence were identified via website databases NexStrain (24), Pango Lineages (25,26) and Centre for Disease Control (CDC) (27). The P.1 variant Spike expression plasmid (pEVAC) was synthesised commercially (GeneArt) with a 19 amino acid C-terminus truncation to increase yields in pseudotyped virus production. The Spike expression plasmids of B.1.1.7 (pI.18), B.1.351 (pI.18) and B.1.1.298 (pcDNA3.1+) were generated in-house by site directed mutagenesis. All plasmids were sequenced to verify successful generation of mutations.
Pseudotype virus generation
We generated pseudotyped viruses (PVs) bearing the Spike protein of the SARS-CoV-2 Wuhan Type and VOCs as previously described (28). Briefly 1000ng of p8.91 HIV Gag-pol, 1500ng of pCSFLW luciferase and 1000ng of SARS-CoV-2 Spike plasmids were resuspended in Opti-MEM and mixed with FuGENE HD (Promega) at a 1:3 ratio. Transfection complexes were then added dropwise in T-75 culture flasks containing HEK293T/17 cells with replenished fresh DMEM at 70% cell confluency. The culture media was harvested 48 hours post transfection and filtered through a 0.45µm cellulose acetate filters. PVs were then titrated and aliquoted for storage at -70°C.
Pseudotype virus titration
The day prior to titration, HEK293T/17 cells were transfected with human ACE-2 (pcDNA 3.1+) and TMPRSS2 (pcDNA 3.1+) expression plasmids using FuGENE HD, to render cells permissible to PVs bearing the SARS-CoV-2 Spike protein. On the day of titration, 100µL of undiluted PV supernatant was serially diluted 2-fold down white F-bottom 96-well plates in 50µL of DMEM. HEK293T/17 cells expressing ACE/TMPRSS2 were added at 10,000 cells per well. Plates were incubated for 48 hours at 37°C and 5% CO2. After incubation, the media was aspirated, and cells were lysed using Bright-Glo reagent (Promega) and luminescence was measured using a GloMax luminometer (Promega). PV entry was measured based on relative luminescence units per ml (RLU/ml).
Neutralisation Assays
Pseudotype neutralisation assays (pMN) were carried out as previously described (28). Briefly, human convalescent plasma was mixed with DMEM at a 1:40 input dilution and serially diluted 2-fold to 1:5,120 in a white F-bottom 96 well plate. PVs were added at a titre around 5×106 RLU/ml in each well. Plates were incubated for one hour at 37°C and 5% CO2, followed by addition of HEK293T/17 cells expressing ACE2/TMPRSS2 at 10,000 cells per well. Plates were incubated for 48 hours prior to assaying with Bright-Glo reagent. Each experiment was performed alongside either the NIBSC 20/162 calibrant and HICC-pool 2, an internal calibrant generated by mixing several serum samples from patients.
Calculation of International Units from IC50 values
IC50 values were calculated for the neutralisation assays based on 4-parameter log-logistic regression dose response curves. These curves were fit using AutoPlate (Palmer et al, under review) and the R package drc (29). Before converting IC50 values into International Units we demonstrated that the assumption of parallel lines was met for different calibrants against each tested variant (Supplementary Figure 1). For each variant we fit two models one allowing each calibrant to have its own IC50 value and its own gradient and one where a single gradient was shared between calibrants. These two models were compared using an ANOVA test.
After demonstrating parallelism between internal calibrants and the WHO International Standard, we calculated the units of our calibrants. The WHO International Standard (NIBSC code 20/136) has a potency of 1000 IU/ml for neutralising antibody activity after reconstitution. We determined the International Units of our internal calibrants against the B.1 virus and VOCs as a ratio of the calibrant’s potency relative to 20/136.
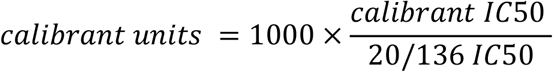 To convert the IC50 of samples to International Units, we calculated the sample’s potency as a ratio relative to that determined for the internal calibrant.
To convert the IC50 of samples to International Units, we calculated the sample’s potency as a ratio relative to that determined for the internal calibrant.
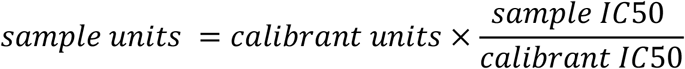 For measurements where the IC50 dilution was less than the minimum tested dilution (1:40) the IC50 value was set to zero. To avoid these samples dominating calculation, one was added to all values when calculating geometric means for IC50 dilutions and International Units.
For measurements where the IC50 dilution was less than the minimum tested dilution (1:40) the IC50 value was set to zero. To avoid these samples dominating calculation, one was added to all values when calculating geometric means for IC50 dilutions and International Units.
International Units allow neutralisation measured in one laboratory against a specific strain to be compared with that measured in a different laboratory. However, it cannot be used to compare neutralisation between different variants.
Statistical methods
Dose response curves were fit to pMN data using AutoPlate software (Palmer et al, under review). To identify escape mutants, we compared how easily different variants could be neutralised by convalescent plasma from patients and previously infected HCWs. Sample potency (IC50) was compared between each variant and the B.1 variant using a paired one-sided Wilcoxon signed rank test in R (30,31). Our one-sided test assumed that the B.1 variant was more potently neutralised than the other VOCs.
We compared neutralisation (IU/ml) by our patient and previously infected HCW cohorts of each VOC using an unpaired test, Wilcoxon rank sum test (30,31). We used a one-sided test which assumed that patients would show greater IC50 values.
We also tested whether the difference in IC50 between patients and previously infected HCWs was the same for different variants. For this we fit a linear mixed model in ‘lme4’ (32) predicting the natural log of the IC50 based on cohort and the variant being neutralised. A random intercept was used to account for measuring each sample against five variants. Only measurements with detectible neutralisation were included in this analysis. After filtering out non-neutralising measurements and log transformation, visual investigation of the residuals showed no trends or violations of the assumption of normality. We also fit a second model with an interaction to allow the effect of cohort to differ between variants. The significance of this interaction was assessed by comparing the two models using an F-test based on the Kenward-Rodger correction (33).
Finally, we investigated how disease severity was related to IC50 in all variants for samples with detectible neutralisation. For this we used a linear mixed model similar to the one described above but using WHO clinical COVID-19 severity scores on the combined group of HCWs and patients.
Results
Neutralisation Responses to circulating SARS-CoV-2 variants
To assess the neutralisation activity of antibodies in convalescent serum from patients (n=38) and previously infected, seropositive HCWs (n=23), pMN were conducted with PVs bearing B.1 Spike and VOCs (Figure 2). IC50 titres showed a significant reduction between B.1 and B.1.1.7 (1.9-fold decrease in geometric mean, p<0.001), indicating that the mutations in the Spike of this VOC does seem to affect antibody neutralisation in the samples tested. B.1.1.298 variant showed a modest decrease in IC50 (1.1-fold decrease, p<0.001) titre, whereas both B.1.351 (8.3-fold decrease, p<0.001) and P.1 variants showed larger decreases (8.2-fold decrease, p<0.001). This data indicates that the Spike protein mutations found in the B.1.351 and P.1 variants are less likely to be neutralised by antibodies generated from previous infection with the B.1 virus isolate.

Neutralisation assays were carried out using pseudotypes expressing either B.1 spike or B.1.1.298, B.1.1.7, B.1.351 and P.1 (A). There were modest decreases in IC50 titres between B.1 and B.1.1.7 (p=<0.001) and B.1.1.298 (p<0.001), followed by a larger decrease in both the B.1.351 (p<0.001) and P1 variants (p<0.001) (B). Points in panel A filled with grey denote patient plasma samples. Wilcoxon signed rank tests were used for statistical analysis between B.1 and each VOC. Black lines in panel A denote geometric means.
Neutralising antibody titres correlate with disease severity
Sub-cohort analysis was used to evaluate antibody titre by disease severity. The results reveal that the patients (n=38) had more potent neutralising antibodies compared to the seropositive HCWs (n=23) (Figure 3) to all variants tested (p<0.001). Patients showed a 16-fold, 16-fold, 10-fold, 21-fold, and 45-fold increase in potency for the B.1, B.1.1.7, B.1.1.298, P.1, and B.1.351 variants respectively. The geometric mean of IC50 values of previously infected HCWs was closest to that of non-infected HCWs (n=36) for B.1.351 and P.1 variants. These data suggest that VOC B.1.351 and P.1 are less sensitive to neutralising antibodies at the lower antibody concentrations known to be found in individuals with a history of asymptomatic infection or mild disease. We used the WHO International Standard to convert all IC50 values into IU/ml to allow for inter-laboratory comparison (Table 2). Due to differing immunoreactivities, each of the variants were independently calibrated to the International Standard.
The geometric means and interquartile ranges obtained from the datasets presented in Figure 3 were converted into International Units (IU/ml) to allow for cross laboratory comparisons. The IU/ml cannot be used to cross compare between variants.
The geometric means and interquartile ranges obtained from the datasets presented in Figure 4A were converted into International Units (IU/ml) to allow for cross laboratory comparisons. Interquartile ranges for datasets in disease severity scores 5 and 6 do not have an interquartile range due to lack of data points.

Neutralisation potencies from different well defined cohorts are presented in IC50 units to allow for comparisons between variants (A) and separate variant graphs with IC50 converted into IU/ml using the 20/136 international standard to permit future cross laboratory comparisons (B) Table representing the geometric means and interquartile ranges in IU/ml to enable cross laboratory comparison. ANOVA tests were used for statistical analysis between the cohort groups. Black lines denote geometric means.
Disease severity correlates with higher IC50 titre across the three VOCs
Finally, we tested whether IC50 was correlated with the WHO clinical criteria of COVID-19 severity and if this relationship was the same for all VOCs (Figure 4B). COVID-19 severity was significantly correlated with IC50, although this relationship did not differ between VOCs (severity F(1, 43.2)=21.0, p<0.001; interaction F(4, 144)=0.60, p=0.66). As before the IC50 values were log transformed. Note that this means we tested proportionate, rather than absolute decrease in neutralising IC50.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Using pseudoviruses expressing B.1 spike, we observed that as severity of COVID-19 increases, the geometric means of nAbs also increases, reaching a plateau at severity scores 4 (severe pneumonia) to 7 (septic shock) (A). Asymptomatic individuals had the lowest titres of nAbs. Symbols filled with grey denote patients, symbols filled in white denote HCWs. To compare IC50 titres from pseudotypes expressing all VOCs spike, IC50 was used as the units of neutralisation as IU/ml does not allow for comparisons against variants (B).
Discussion
The COVID-19 pandemic has resulted in multiple nationwide lockdowns and renewed efforts to accelerate vaccination programs around the globe, there is a strong urge to return to normality to mitigate further damage to livelihood and economies. Unfortunately, the political urge to prematurely lift public health and social restrictions has resulted in large surges and new waves of regional epidemics while national immunisation programmes are in early stages and population level immunity is incomplete. While there is growing evidence that in the few countries with progressive immunisation programmes there is currently decreasing clinical cases and hospitalisation, relieving pressure on health care infrastructure, there is concern that the VOCs may continue to circulate and evolve resistance to vaccine induced immunity. A recent report from Israel alarmingly found an increased incidence of the B.1.351 variant amongst vaccinees (34). As a result, understanding the role of pre-existing natural immunity in reducing disease severity is a key factor for informing policies of governments eager to reopen their economies.
The emergence of variants has become a significant issue that needs to be taken into consideration. The B.1.1.298 variant contains a unique mutation, Y453F, in the RBD, which was found to enhance ACE2 binding affinity(35). Y453F has not been detected in any of the other variants mentioned in this article, though two new variants in circulation, B.1.429 emerged in California, USA and B.1.617 which emerged from India and just recently categorised as a VOC by the UK Government, contains an adjacent mutation at position L452R which may also impact antibody neutralisation (36–38). B.1.1.298 also includes a deletion at amino acids positions 69 and 70 (69/70del). This particular deletion in the N-terminus of Spike was suggested to affect the structure of the Spike in a way that may facilitate immune evasion(39), and has also been detected in the B.1.1.7 variant. There are several reports that showed a decrease in neutralisation capabilities of antibodies generated by either B.1 infection or vaccination against the B.1.1.298 variant, which is consistent with our data. (40,41). Nevertheless, due to the massive culling of mink in farms in Denmark, it is widely believed that the incidence of spillover of this variant from mink to humans has been largely eliminated. The B.1.1.7 variant, which arose in the southeast of England after the first wave of B.1, is currently the most prevalent VOC in circulation in the UK. The clinical significance of this is uncertain (41–44). Several studies have investigated whether B.1.1.7 escapes immune evasion from antibodies generated by vaccination (45,46). One study observed a decrease in antibody neutralisation titres against B.1.1.7 from individuals who had been vaccinated with a single dose of BNT162b2 mRNA vaccine, resulting in a 3-fold reduction of IC50, though this drop in neutralisation was less evident after the second dose of vaccination (46). An independent study found similar results with the both BNT162b2, ADZ1222 vaccine and convalescent serum from infected patients (45).
The largest reduction in neutralising antibody titres were with P.1 and B.1.351 variants, both of which have the E484K mutation. The impact of E484K mutation on immune evasion has been reported in multiple studies (9,12,41,42,47,48). The immune evasion displayed by P.1 and B.1.351 is concerning. These two VOCs pose a risk to individuals who have had an infection, that may potentially lead to reinfection cases, as has been reported recently in 3 patients from Brazil with respect to the P.1 variant (11,13,18), and may disproportionately affect those vaccinated compared with those unvaccinated in the same community (34).
Standardisation in the reporting of data is critical for comparison of data in different populations and countries and to harmonise assay to assay and lab to lab variability, which will be vital in informing national and international public health policies around the world (49). Here we report these findings in International Units established by the use of WHO International Standard comprising a pool of 11 convalescent plasma sourced from the first wave of global infections established with relative genetically homogeneous SARS-CoV-2. One of the main questions regarding the antiviral neutralising antibody responses remains: what are the immune correlates of protection against SARS-CoV-2? Some studies have investigated immune correlates of protection in both animal and human models (50), but an issue remains with the units of antibody titres. Due to laboratory variations in experimental procedures, IC50 values cannot be directly compared across multiple studies (22). To overcome this issue, the WHO International Standard, which has been analysed in a large collaborative study including 51 laboratories and multiple serological tests, allows for standardisation of assay results and cross laboratory comparisons(22). Our use of the WHO International Standard has permitted us to convert IC50 values into International Units. As a result, our data can be compared with other laboratories that report data in International Units. Importantly, the WHO International Standard cannot be used to compare against the different immunoreactivities of variants and thus calibration must be carried out for each separate variant, and data reporting, must specify the variant analysed, which then allows for multiple laboratories to compare datasets against the same variant. Crucially, data converted into IU/ml cannot be used to compare across different variants. We have reported differences between cohorts against the same variant in International Units but when comparisons between variants were assessed, we used IC50. Continuous accumulation of neutralisation data from multiple laboratories represented in International Units could greatly assist in determining immune correlates of protection, which would be deeply informative in the progress of understanding SARS-CoV-2. An example of this would be to determine the minimum antibody titre required for protection. This could be feasible where disease severity scores are known for each source of plasma or serum that was tested by neutralisation assay. By extension, identifying the minimum titres that constitute protection could lead to greater knowledge of whether a novel vaccine would be considered protective against the B.1 virus.
In summary, this data, expressed in IU/ml, represents a benchmark “pre-vaccine” standardised dataset comparing infected individuals with different disease outcomes. This will allow multiple laboratories to compare IC50 values in IU/ml with this data providing their assays are calibrated against the WHO International Standard for each studied variant, without attempts to use IU/ml to compare IC50 values across any current and future variants. The continual use of the Standard by various laboratories could greatly increase our ability to establish benchmarks, or thresholds of correlates of immunity. The next steps involve expanding this standardised data to immunised individuals for comparison of neutralising antibodies in convalescent, versus infected and vaccinated individuals against the different VOCs and establishing thresholds of protection against circulating variants to inform national and international vaccine programmes.
Data Availability
Raw data will made available upon request.
Acknowledgements
This study was undertaken by the Humoral Immune Correlates to COVID-19 (HICC) consortium, funded by the UKRI and NIHR; grant number G107217 (COV0170 - HICC: Humoral Immune Correlates for COVID19). RW and SE received funding from the StMWK (ForCOVID, Bavaria, Germany). We thank the RPH Foundation Trust COVID-19 Research and Clinical teams for supporting recruitment to this study, HCWs and Outpatients who participated in this study.
Footnotes
↵8 Humoral Immune Correlates for COVID-19 Consortium
References




