
Abstract
Alpha-synuclein pathology is associated with immune activation and neurodegeneration in Parkinson’s disease. The immune activation involves not only microglia but also peripheral immune cells, such as mononuclear phagocytes found in blood and infiltrated in the brain. Understanding peripheral immune involvement is essential for developing immunomodulatory treatment. Therefore, we aimed to study circulating mononuclear phagocytes in early- and late-stage Parkinson’s disease, defined by a disease duration of less or more than 5 years, respectively, and analyze their association with clinical phenotypes. We performed a cross-sectional multi-color flow cytometry study on 78 gender-balanced individuals sporadic Parkinson’s disease, 28 controls, and longitudinal samples from seven patients and one control. Cell frequencies and surface marker expressions on natural killer cells, monocyte subtypes, and dendritic cells were compared between groups and correlated with standardized clinical scores. We found elevated frequencies and surface levels of migration-(CCR2, CD11b) and phagocytosis-(CD163) markers, particularly on classical and intermediate monocytes in early Parkinson’s disease. HLA-DR expression was increased in advanced stages of the disease whereas TLR4 expression was decreased in women with Parkinson’s Disease. The disease-associated immune changes on CCR2 and CD11b correlated with worse cognition. Increased TLR2 expression was related to worse motor symptoms. In conclusion, our data highlights the TLR2 relevance in the symptomatic motor presentation of the disease and a role for peripheral CD163+ and migration-competent monocytes in Parkinson’s disease cognitive defects. Our study suggests that the peripheral immune system is dynamically altered in Parkinson’s disease stages and directly related to both symptoms and the sex bias of the disease.
Highlights
TLR2 expression increased in patients worse motor symptoms.
Increased CD163 and HLA-DR monocytic expression in patients with long PD duration.
Sexual-dimorphism for CCR2, CD11b, and TLR4 expression on PD monocytes.
CCR2 and CD11b expression are associated with cognitive impairment in PD.
1. Introduction
The immune component of Parkinson’s disease (PD) has been settled by studies showing microglia activation in post-mortem brains (Imamura et al., 2003a), low-level systemic inflammation (Qin et al., 2016), and decreased PD risk in NSAID users (Gagne and Power, 2010; Ren et al., 2018). Moreover, variants in immune-related genes, including HLA-DR, are associated with increased PD risk (International Parkinson Disease Genomics et al., 2011; Rayaprolu et al., 2013), which suggests a significant role for both the innate and the adaptive immune system. Accordingly, T-cells from people with PD (PwP) exhibit a Th1/Th17 bias (Storelli et al., 2019), and they respond to α-synuclein (α-syn)-derived peptides early in PD (Lindestam Arlehamn et al., 2020; Sulzer et al., 2017). Despite the importance of the innate immune system’s role in activating/priming the adaptive system, there is a paucity of evidence on the innate peripheral immune cells’ changes in PD. Most research has focused on microglia, however, peripheral immune cells infiltrate the brain and participate in the inflammation and neurodegeneration in PD (Harms et al., 2021). With increasing evidence that PD is not only a CNS disorder but also affects and, in some cases, originates in the periphery (Horsager et al., 2020), more knowledge is required regarding the peripheral innate immune cells including natural killer (NK) cells and peripheral mononuclear phagocytes (MNPs).
We have previously shown that soluble (s)CD163, a marker produced upon activation of CD163-expressing MNPs, was increased in PD biofluids (Nissen et al., 2021). This elevation was related to PD-stages, defined by the disease duration, and associated with cognition and neuronal disease biomarkers such as α-syn and Tau. This suggests an active involvement of MNPs in the different stages of PD. MNPs comprise populations of ontogenic, phenotypic, and functionally overlapping monocytes (Mos) and dendritic cells (DCs) (Dutertre et al., 2019; Geissmann et al., 2003). MNPs act as myeloid precursors that enter the tissue and differentiate into Mo-derived macrophages (Mϕs) or DCs, (Dutertre et al., 2019; Geissmann et al., 2003). Daily, half of the circulating MNPs leave the bloodstream under steady-state conditions to infiltrate tissue, and this can increase during inflammation (Geissmann et al., 2003). Based on their expression of CD14 and CD16, Mos are typically subdivided into classical (cMo, CD14high/CD16-), intermediate (iMo, CD14low/high/CD16+), and non-classical (ncMo, CD14-/low/CD16++) (Abeles et al., 2012). cMos, and to some degree also iMos, migrate to inflamed tissue in a CCR2-dependent manner and are highly phagocytic. In contrast, ncMos are described as blood-endothelia patrolling cells migrating to non-inflamed tissue, including the brain, in a CX3CR1-dependent manner (Geissmann et al., 2003). MNPs also include a fraction of the dendritic cells (DCs, CD14-/low/CD16-), which are responsible for antigen presentation to and priming of T-cells. Although DCs in general do not express CD14 and CD16, they share many other surface receptors with Mos, such as TLR2, HLA-DR, CCR2, and CD163. Infiltration of CCR2+ or CD163+ cells seem to partake in the inflammatory and neurodegenerative processes in PD brains (Harms et al., 2018; Tentillier et al., 2016).
Immune changes in the brain (Edison et al., 2013) associated with MNPs are suggested to be highly relevant in PwP with significant cognitive decline (Nissen et al., 2021; Wijeyekoon et al., 2020a). Interestingly, the presentation of PD differs between sexes (Iwaki et al., 2021), with cognitive impairment being less likely to occur in female patients. We have previously reported a sex dimorphism in the CD163 related immune changes in serum from PwP (Nissen et al., 2021). In addition, it has recently been shown by transcriptomic analysis of blood CD14+ Mos that female with PD show a homogenous monocytic response while the response was more heterogenous in males (Carlisle et al., 2021). All this supports a sex-specific immune response that might be associated with the differences in disease presentation, especially with regards to cognition. To study the profile of peripheral MNPs ex vivo in PD, we used multicolor flow cytometry on a cross-sectional cohort of patients with sporadic PD and seven longitudinal PD cases. Based on our prior data, we divided the patients in the cohort in two groups: those whose samples were collected early after disease diagnose (early PD) and those collected late in the disease (late PD) using five years as a cut-off, and we analyzed putative sex differences. The data was then correlated to motor and non-motor symptoms.
2. Materials and methods
2.1 Cohorts and sample collection
Patients with sporadic PD, diagnosed according to the UK Brain Bank criteria, and healthy controls (HCs) with no signs of neurodegenerative diseases were recruited at the University Hospital Tuebingen, as previously described (Goldeck et al., 2016; Hughes et al., 1992; Nissen et al., 2019) (Supp.Fig.1). The local ethics committee approved the study (480/2015BO2), with all participants providing written informed consent.
Peripheral blood mononuclear cells (PBMCs) were collected, isolated, and stored at the Hertie Neuro-Biobank (University of Tuebingen) and analyzed at Aarhus University (see supplementary information). The study includes: Cohort#1A: Cross-sectional samples from 78 PwP collected early (< 5 years) or late (≥5 years) after PD diagnose and 28 HCs (Table 1); and Cohort#1B: Longitudinal samples from seven PwP and one HC (Supp.Table 1). Information on comorbidities and pharmacological treatment for Cohort#1A is provided in Supp.Table 2. From the samples received (n=109), three individuals with known autoimmune comorbidities (AID) were excluded: 1 female control and 2 late PD, one female and one male (Supp. Fig.1).
Cohort#1A for cross-sectional analysis of blood samples collected from healthy control (HC) and people with sporadic Parkinson’s disease (PD) early (<5 years, EPD) or late (≥5 years, LPD) after diagnose, without known autoimmune comorbidities, and with an equal distribution between females (F) and males (M). Mean, standard derivation (SD), and the number (N) of individuals for whom the data were available from at the sampling date are shown for: L-dopa equivalent daily dose (LEDD), Hohn and Yahr (H&Y), Unified Parkinson’s Disease Rating Scale three (UPDRS III), the Montreal Cognitive Assessment (MoCA) score, Beck Depression Inventory II (BDI II), and Sniffin’ Stick 12 olfaction test (SS12). Storage time for the frozen samples in each group was similar. ¤/* parametric and non-parametric significant p values are shown the corresponding unpaired t-tests or one-way ANOVA with multiple comparison: * p<0.05, ** p<0.01, *** p>0.001, **** p<0.0001.
2.2 Flow cytometry
PBMCs were stained for flow cytometry with a combination of nine fluorochrome-conjugated antibodies (Supp.Table 2), as previously described (Farmen et al., 2021). The cells were run on an LSR Fortessa (BD) flow cytometer and data were analyzed using FlowJO V10. For details, see supplementary information.
2.3 Statistical analyses
GraphPad Prism V7 was used for statistical analyses. In Cohort#1A, normality was informed with D’Agostino-Pearson omnibus test. Sex differences were investigated for each analysis, if no significant differences were observed data from both sexes were merged. Group differences were analyzed using One-way ANOVA (or Kruskal-Wallis test) with Tukey’s (or Dunn’s) post hoc analysis. Associations between markers and clinical data were tested using Pearson (or Spearman) correlations as appropriate with two-tailed p values, and if significant assessed for relevant covariants using simple and multiple linear regressions by JMP. Only associations with p<0.05 for both analyses (correlation and linear regression) were considered, unless indicated. Ordinary 2-way ANOVA was used to investigate related populations with dependent gating.
For comparison of longitudinal sampling at different months in Cohort#1B, normality was assumed and paired repeated-measures mixed-effect model (REML) with Tukey’s multiple comparisons performed. The first and last visit were compared with non-parametric paired t-tests (Wilcoxon).
We evaluated the effect of anti-inflammatory drugs and found no difference in the mean of each group including individual with such treatment and without. Furthermore, the values of these individuals were evenly spread. Therefore they were included in the analysis.
3. Results
3.1 Demographics
After AID exclusion, Cohort#1A included sporadic 78 PD patients and 28 HCs with similar sex-(50%) and age-distribution (HC 64.2±7.4 & PD 66.6±9.5; mean±SD). We subdivided the PD patients in two groups according to the disease duration at sampling: early (<5y) or late PD (≥5 years) based on our prior data on soluble (s)CD163, which supported a stage-associated (disease duration) increased MNP activation in PwP (Nissen et al., 2021). Equally, that data also indicated a sex difference in the MNP activation in patients’ blood; thus, sex differences were interrogated for each analysis. This subdivision resulted in a late PD group significantly older than the early PD group and HCs, which was also significant when investigating the males only, not females (Table 1). Levodopa equivalent daily dosage (LEDD) was higher in late vs. early PD, and also higher in late males vs. late females (Table 1). Thus, all differences found among these groups were appropriately compensated for the relevant covariant.
Additional eight individuals provided longitudinal samples: Cohort#1B. Two to four samples were available per individual, with a timeframe of one to sixteen months from the first to the last visit (Supp.Table 1). Cohort#1B was analyzed separately.
3.2 Unraveling the MNPs in the PBMC pool by TLR2 expression
Classical gating of Mos by forward- and side scatter prior to CD14/CD16 subtyping, results in an ignored fraction also including circulating DCs (Geissmann et al., 2003). Instead, we used CD56 and TLR2 as inclusion markers for natural killer (NK) cells and MNPs, respectively. MNP (TLR2+) included all Mos and some DCs, as shown by tSNE plot of concatenated samples of live cells (Supp.Fig.2): All CD14+ and CD16+, except the CD16+/CD56+ cells, were TLR2+. Hence, our gating distinguishes the (TLR2+) MNPs from the (TLR2-/CD56dim) mature- and (TLR2-/CD56bright) precursor NK cells, from the remaining PBMC pool (TLR2-/CD56-) (Supp.Fig.3).
The TLR2+ MNPs were divided into four subpopulations based on CD14/CD16 and HLA-DR expression (other characteristics in brackets): cMos CD14++/CD16-/HLA-DRlow (CD11b+/CCR2bright/CD163bright/TLR2bright), iMos CD14++/CD16+/HLA-DRbright (/CD11b+/CCR2bright/CD163bright/TLR2bright), ncMos CD14dim/CD16++/HLA-DRdim (/CD11b- /dim/CCR2+/CD163dim/-/TLR2bright), and the DCs CD14dim/-/CD16-/HLA-DRbright (/CD11b- /CCR2+/CD163Mixed/TLR2dim) (Supp.Fig.3).
In-depth analysis of DCs discriminates the plasmacytoid (p)DC and the antigen-presenting conventional (c)DC type 1 and 2. An additional antibody panel on three control donors (not included in the cohort#1A nor #1B) confirmed that the CD1c+ cDC2s are TLR2+, thus, included in our MNP population, whereas both the CD141+ cDC1s and the CD303+ pDCs (Dutertre et al., 2019) were TLR2- (Supp.Fig.4). The cDC2 is responsible for Th1/Th17 CD4 T-cell priming (Dutertre et al., 2019). We showed that the TLR2+ cDC2s primarily were located in the DC gate being CD14-/dim/CD16-, with a smaller CD14++/CD16- fraction, thus in the cMos gate (Supp.Fig.4). With our gating, cDC2s constituted ∼5% of the MNPs, with ∼90% of the CD14-/CD16- MNPs being cDC2s. For simplicity, the TLR2+/CD14low/CD16- are referred onward as DCs.
The percentage of viable cells in the flow staining was increased in late PD, although this did not remain significant after adjusting for age (p=0.052) (Supp.Fig.5A). Nevertheless, PBMC viability correlated negatively with the Montreal Cognitive Assessment (MoCA) scores for all PwP, even when adjusted for age and LEDD (Supp.Fig.5B, not analyzed in HC due to only five MoCA scores informed in the group). Hence, worse cognition was associated with better PBMC viability. Accordingly, when dividing the cohort according to MoCA score (>26=no cognitive impairment (noCI) and ≤26=cognitive impairment (CI)), we found a significant increase in cell viability for the CI group (Supp.Fig.5C).
3.3 Mature NK cells in PwP are activated and primed for antibody-dependent cytotoxicity
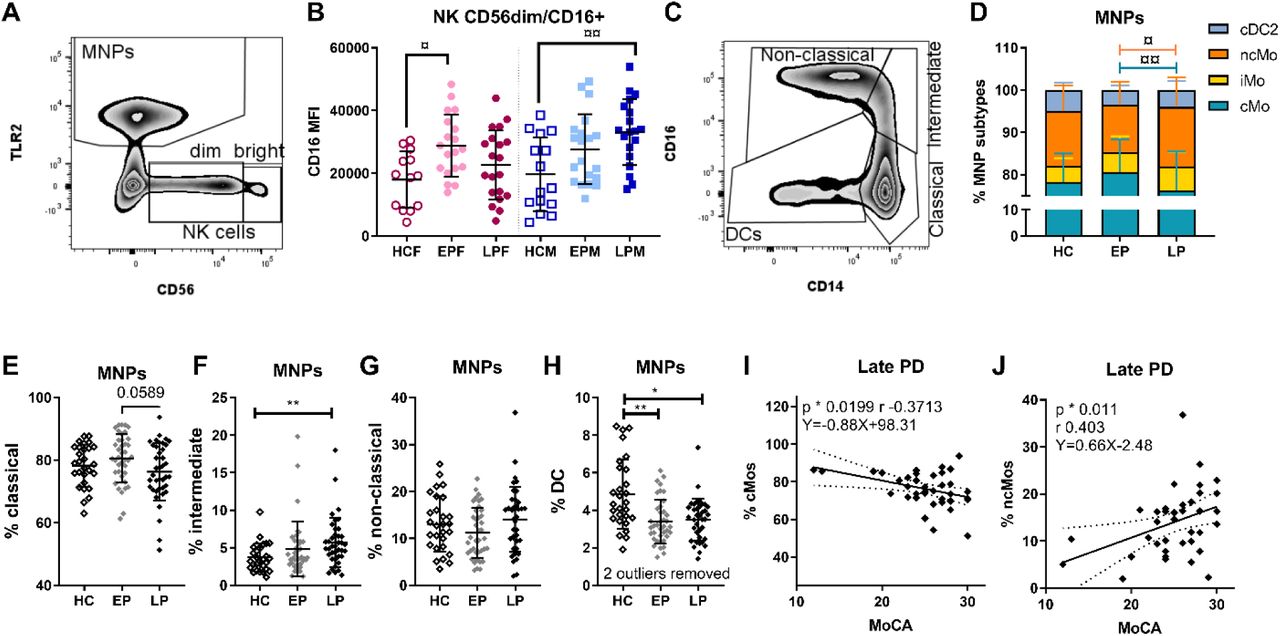
Peripheral NK cells migrate to the CNS during synucleinopathies (Earls et al., 2020). Based on CD56 expression we separated NK cells into precursor and mature population. Since CD16 increase on NK cells has been related to activation of NK cells (Earls and Lee, 2020b), we also analyzed the activated CD16+ fraction within the mature NK population (Fig.1A-B). We observed an increase in CD16 median florescence intensity (MFI), i.e., upregulation of the antibody receptor FCγRIII expression, on the mature NK cells during early PD for females and late PD for males (Fig.1B), which remained after compensating for age (the significance was lost for females when also adjusting for LEDD).

A) Gate for identification of all mononuclear phagocytes (MNPs) and natural killer (NK) cells divided into mature (CD56dim) and precursor (CD56bright) populations. B) CD16 median fluorescence intensity (MFI) on CD16+ cells within the CD56dim mature NK population in healthy controls (HC), early PD (EP), and late PD (LP) in female (F) and male (M) groups. C) Gating of MNP subtypes and D) comparison of the distribution of MNP subtypes (¤/¤¤ 2-way ANOVA with Tukey’s test multiple comparisons adjusted p values). Frequency of monocyte subtypes E) classical (cMo), F) intermediate (iMo), G) non-classical (ncMo), and H) conventional dendritic cell type 2 (cDC2) of the total MNP population. For the late PD group, Pearson correlations (p & r) and linear regressions (equation) between Montreal Cognitive Assessment (MoCA) score and the frequency of I) cMos and J) ncMos. Statistical approach E-H): */** non- parametric 1-way ANOVA, cMos: parametric 1-way ANOVA with multiple comparisons. */¤ p <0.05, **/¤¤. p <0.01. E-H are interrelated based on the gating; thus, the p values were Bonferroni corrected for multiple-comparison: all */** p values remained below the adjusted threshold of 0.0125.
3.4 PD stage-related changes in the distribution of the MNP subtypes
The MNP percentage among PBMCs was similar for: HCs (mean±SD: 23.25%±6.60), early (27.01%±8.68), and late PD (24.22%±10.53) and it was not affected by stage or sex. However, the MNP subtypes distribution was altered by the disease stage (Fig.1C-I). A 2-way ANOVA analysis showed a significant interaction between subtypes and disease stages (F(6, 412) = 3.615, p=0.0017), and a post hoc analysis for multiple comparisons by Tukey’s test revealed a significant decrease of the cMos from early to late PD and a mirrored increase of ncMos in late compared to early PD (Fig.1D). Individual MNP subtype analyses with Bonferroni correction (alfa 1.25%) (Fig.1E-H), showed significant increase of iMos in late PD vs. HC (Fig.1F), which was reflected in a significant drop of DCs in early and late PD vs. HC (Fig.1H) that remained after compensating for age. Notably, we observe a significant, although weak, positive correlation between the percentages of DCs with the Sniffin’ sticks 12 olfaction scores in PwP (Supp.Fig.5D). Hence, fewer DCs were associated with hyposmia, which was not driven by covariants. Furthermore, in the late PD cases, the fraction of cMos and ncMos correlated negatively and positively with MoCA, respectively (Fig.1I-J). Hence, those patients where the number of cMos in blood did not decrease back to healthy levels at late PD (and vice versa with ncMos), were those developing cognitive impairment.
3.5 Altered expression of receptors responsible for α-syn sensing and presentation
Variants in the gene encoding HLA-DR are associated with PD and the molecule is responsible for presenting α-syn to adaptive immune cells (Harms et al., 2021) hence we investigated its expression in the cohort. The Mo populations expressing high level of HLA-DR, the iMos and ncMos, showed increased HLA-DR expression (MFI) in PwP (HC vs. PD, iMos p=0.01 and ncMos p=0.013, non- parametric t-tests), although when separated in stages this was only significant at late PD (Fig.2A-B). This remained significance when adjusting for age. However, these late increases correlated with LEDD and the significance was lost when compensated for L-DOPA dose.
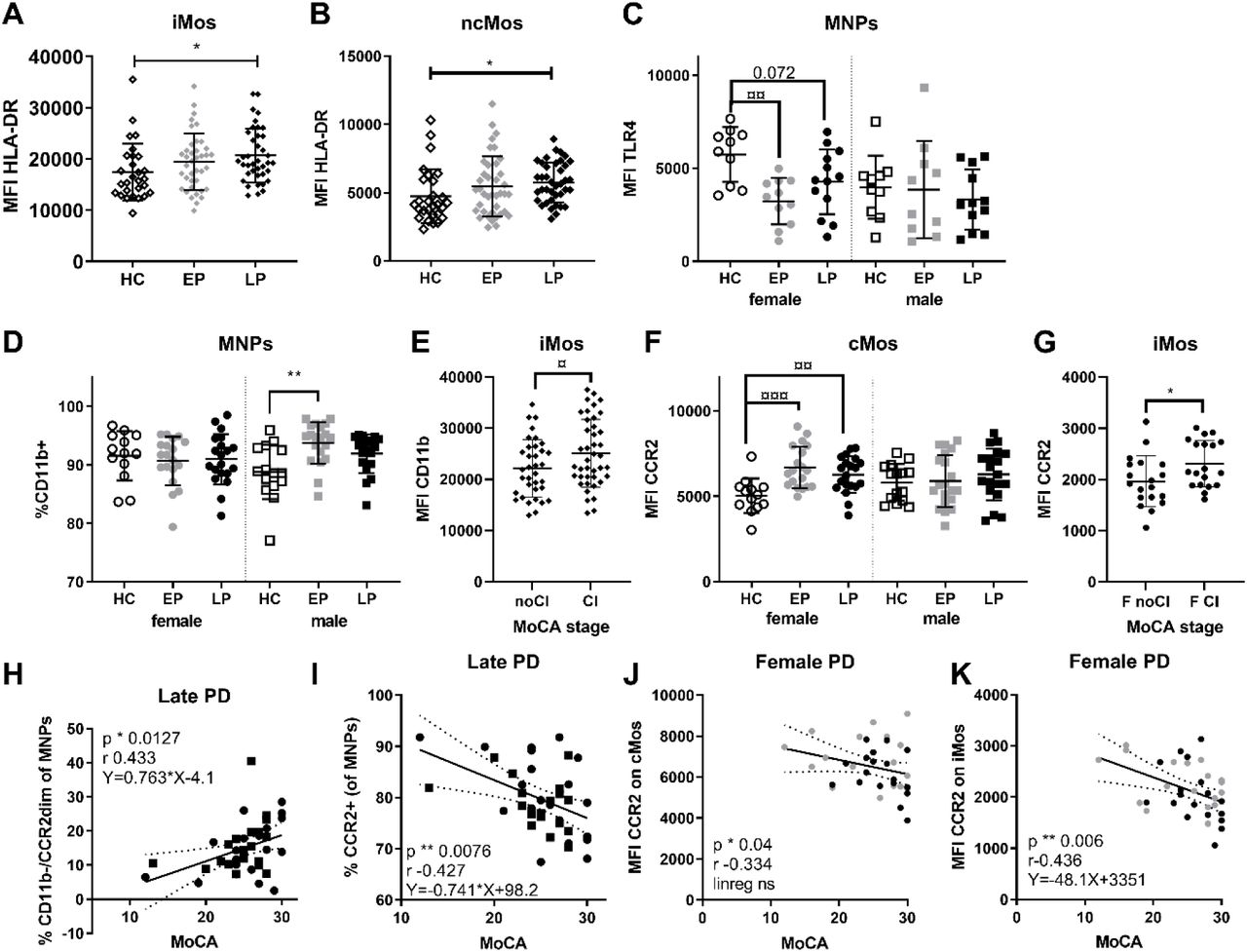
Median fluorescence intensity (MFI) of HLA-DR on A) intermediate (iMos) and B) non-classical monocytes (ncMos) in healthy control (HC), early PD (EP), and late PD (LP) groups. C) MFI of TLR4 on all mononuclear phagocytes (MNPs); and D) the percentage of CD11b+ MNPs, analyzed sex separated. E) The MFI of CD11b on iMos in patients with no cognitive impairment (noCI), i.e. Montreal Cognitive Assessment (MoCA) scores >26; or with cognitive impairment (CI, MoCA≤26). F) MFI of CCR2 on cMos analyzed by group and sex; G) and CCR2 MFI on iMos in females (F) with or without cognitive impairment (CI vs. noCI). MoCA scores in patients in the late PD group correlated H) positively (p=0.041 after correction for age covariance) with the fraction of CD11b-/CCR2dim MNPs (identified in Supp.Fig.3F); and I) negatively with the total fraction of CCR2+ MNPs, (one outlier removed using Grubs, p=0.035 after correction for age covariance). In all females with PD, MoCA was negatively correlated with MFI of CCR2 on J) classical monocytes (cMos) and K) iMos, but only iMos showed also a significant linear regression that remained after adjusting for age (p=0.03). Statistics: non-parametric/parametric one-way ANOVA tests with appropriate multiple comparison: */¤ p <0.05, **/¤¤ p <0.01, ***/¤¤¤ p <0.001. Mean with SD, Pearson correlation p and r values, and linear regression equation with 95% confidence intervals are shown. Uncorrected p values are shown.
We analyzed the innate sensors TLR4, which can recognize α-syn and can also facilitate α-syn clearance {(Harms et al., 2021). Our TLR4 analyses only included 66 samples due to antibody lot number-associated variance. We observed lower TLR4 MFI on MNPs in males vs. female HCs, thus, sexes were separated (Fig.2C). This revealed a reduced TLR4 expression on females’ MNPs, particularly in early PD that remained after covariance adjustment. Thus, HC females had TLR4 levels higher than PD females and all males (observed across all MNP subtypes, not shown).
3.6 Sex-dependent changes related to MNP migratory capacity
We examined the integrin CD11b (Fig.2D-E) and the chemokine receptor CCR2 (Fig.2F-G), molecules important for MNP’s migration. Although the percentage of CD11b+ MNPs was similar among sexes in HCs, the percentage of CD11b+ MNPs were increased in early PD males vs. HC, but not in females (Fig.2D). This passed the covariance tests. The MFI of CD11b did not show any apparent PD-related changes, however, when examine the MFI of CD11b on iMos we observed a significant increase in patients with cognitive impairment vs. those without any cognitive impairment according to MoCA scores (Fig.2E)
Regarding CCR2, another migration marker highly expressed on cMo and iMo, we observed increased expression on cMos in (particularly early) female patients (Fig.2F) that remained after compensating for age and LEDD; while no significant differences were seen in males. The iMos also showed a significant upregulation in the MFI of CCR2 predominantly in female patients with cognitive impairment (Fig.2G). This suggests mobilization of the Mos to inflamed tissue, associated with CD11b in male patients and CCR2 in females.
Interestingly, in late PD, the percentage of MNPs with no/low expression of CD11b/CCR2, hence less transmigration capacity, correlated positively with better cognition score in both sexes. Conversely, the percentage of CCR2bright MNPs correlated negatively with cognition (Fig.2H-I; both remained after adjusting for age, but the first missed significance after also adjusting for LEDD p=0.062). Furthermore, in females only, the expression of CCR2 on cMos and iMos also had a negative correlation with MoCA (Fig.2J-K); for the iMos the linear regression was also significant even when age and LEDD-adjusted. Overall, changes of markers related to Mo migration were associated with cognitive symptoms, with higher migratory capacity correlated to worse cognition.
3.7 Peripheral CD163 changes distinguish PwP from HCs
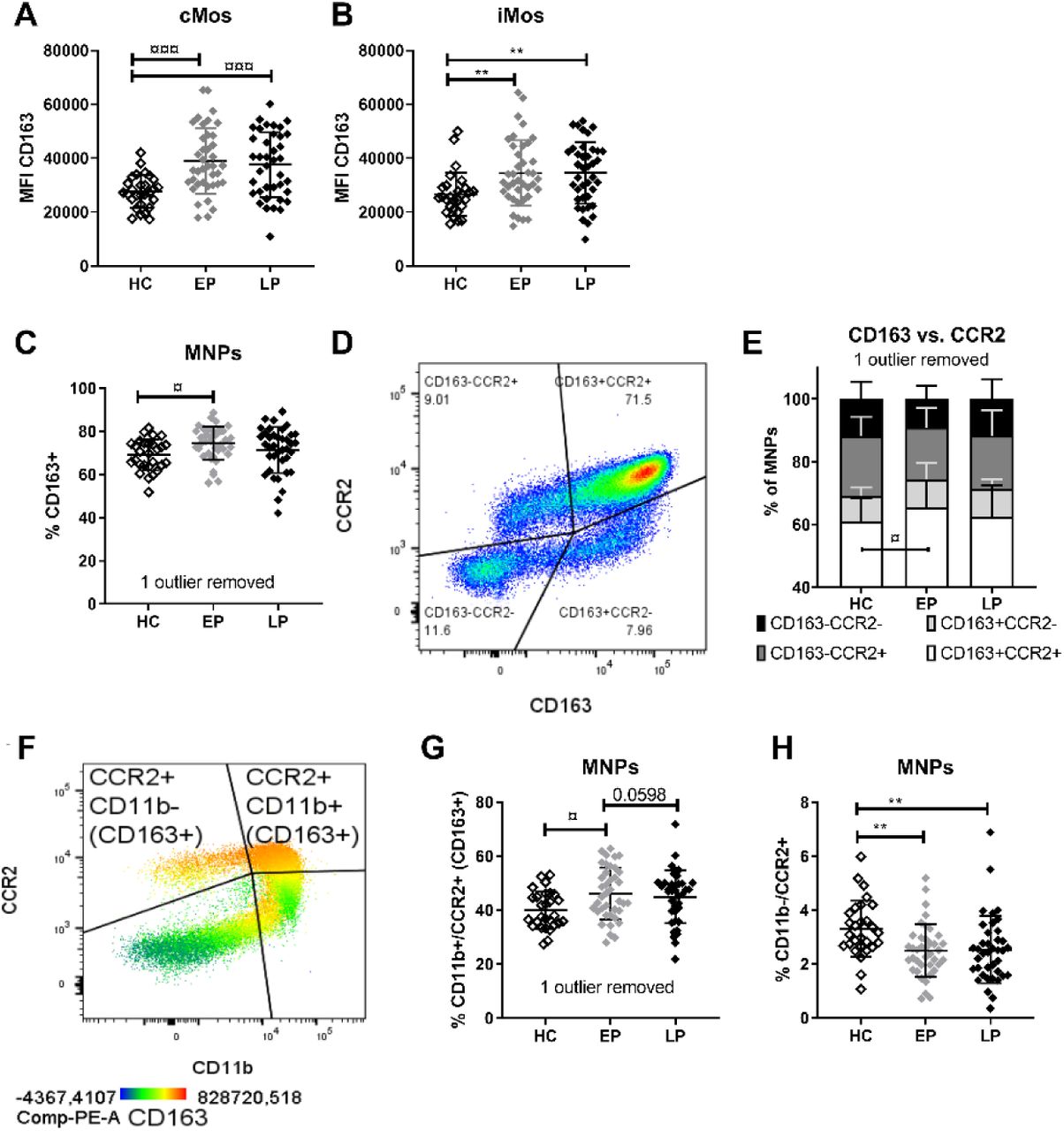
CD163 is a MNP specific protein whose cleavage from the membrane surface is associated with Mo activation and production of sCD163 (Moller, 2012). We have previously shown that PwP’s blood immune cells in vitro have reduced ability to downregulate CD163 expression on CD14+ Mos upon exposure to α-syn (Nissen et al., 2019); and that PwP have increased sCD163 levels in serum (females only) and in CSF (both sexes), with CSF sCD163 levels associated with α-syn and cognitive deficits (Nissen et al., 2021). This data suggested a role for CD163+ cells in PD, thus we here analyzed the CD163 expression on MNPs. Interestingly, PwP showed higher CD163 expression (MFI) on the high expressing cMo and iMo compartments (Fig.3A-B) that passed covariance testing. Furthermore, we observed an increased frequency of CD163+ MNPs in early PD (Fig.3D) that remained after adjusting for age and LEDD.

Median fluorescents intensity (MFI) of CD163 on A) classical (cMos) and B) intermediate (iMos) monocytes. C) Frequency of CD163+ mononuclear phagocytes (MNPs). D) MNPs were gated according their CD163 and CCR2 expression and E) the distribution of the four subpopulations in the groups was compared with 2-way ANOVA followed by Tukey’s multiple comparison test. F) MNPs analyzed based on CCR2 and CD11b, with the majority of CCR2+ MNPs, being also CD163+. The G) CD11b+/CCR2+ fraction, and H) the frequency of CCR2+/CD11b- cells are compared among groups (with Bonferroni corrected p=0.0125 threshold). For simplicity, in this figure, we referred to CCR2 as -/+ populations, but for the MNPs it is in fact dim/bright populations. Adjusted multiple comparisons p values: non-parametric/parametric: */¤ <0.05, **/¤¤ <0.01, ***/¤¤¤ <0.001, ****/¤¤¤¤ <0.0001. Healthy control (HC), early PD (EP), late PD (LP) groups.
Since CD163+ infiltrating cells have been observed in PD models and post-mortem brains of PwP (Harms et al., 2017; Pey et al., 2014b; Tentillier et al., 2016), we investigated the phagocytic CD163+ cells with respect to CCR2 co-expression, regardless of MNP subtyping (Fig.3D/Supp.Fig.3E). A two-way ANOVA analysis (F(6, 408) = 2652, p=0.016) and a post hoc Tukey’s multiple comparisons analysis revealed a significant stage-related change of CD163+/CCR2+ MNPs, which increased in early PD compared to HCs (Fig.3E). We then examined the CD11b+/CCR2+ MNPs co-expressing both migration markers, of which the majority also are CD163+ (Fig.3F/Supp.Fig.3F). MNPs from early PD had more “triple-positive” cells compared to HCs (and a trend vs. late PD p=0.059) (Fig.3G). This remained after covariance adjustment, even though LEDD also had a significant relation. Conversely, both early and late PD showed fewer CCR2+ MNPs lacking CD11b (CD11b-/CCR2+) vs. HCs (Fig.3H) that remained significant after covariance adjustment. Altogether, PwP showed an increased expression of CD163 in MNPs at all stages, and an early expansion of the CD163+ cells co-expressing CCR2 and CD11b.
3.8 TLR2 expression increases longitudinally and with H&Y (Cohort#1A and #1B)
The analysis of the eight longitudinal samples (Cohort#1B) revealed a progressive MFI increase of the oligomeric α-syn receptor TLR2 on total MNPs and cMos alone (first vs. last visit, male PD) (Supp.Fig.6A-B). The TLR2 MFI on ncMos was also significantly increased from baseline sampling to 9-12 months later (Supp.Fig.6D). These TLR2 MFI changes were also reflected in the cross-sectional Cohort#1A when comparing PwP of H&Y ≤2 vs. >2 (Fig.4), showing increased TLR2 MFI on iMos, ncMos, and DCs. Thus, increased TLR2 on MNPs is associated with disease progression and motor worsening. This was not an artefact due to storage time, since TLR2 MFI was not related to samples’ freezer storage time (not shown).

Median fluorescents intensity (MFI) of TLR2 on A) classical (cMos), B) intermediate (iMos), and C) non-classical (ncMos) monocytes as well as D) dendritic cells (DCs) in patients with Hohn and Yahr (H&Y) scores ≤2 versus those with H&Y >2. Non-parametric unpaired t-test, ** p<0.01.
4. Discussion
This study examined MNP and NK cell frequencies and their expression of relevant markers in PwP at early- and late-stages since the PD diagnose in a cross-sectional cohort (#1A). We observed that PD is associated with significant alterations in CD163 expression, migration markers (CCR2 and CD11b), and the HLA-DR. Furthermore, PwP showed stage-related alteration in the frequency of MNP subtypes and receptors compared to HCs (summarized in Fig.5). Some changes in the MNP population were sex-specific or were correlated with PD symptoms, supporting an association between peripheral innate immune cells and disease pathology. More specifically, we found that the disease was associated with an increase in iMos in late stages and an early and long-lasting decrease of DCs with fewer DCs correlated with hyposmia. When comparing among PD stages, we observed enriched cMos in early vs. late PD, with a mirrored expansion of ncMo in late PD. Notably, these alterations were related to the increased frequency and expression of the CD163 scavenger receptor and the migration markers CCR2 and CD11b, especially in early PD. While increased HLA-DR expression was more pronounced in late PD. CD16 expression on mature NK cells was also increased in PwP, with a different temporal development according to sex. The only downregulated immune marker in this PD cohort was TLR4, decreasing in females with early PD (Fig.5).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Summary of study findings based on flow cytometry on peripheral mononuclear cells (PBMCs) from people with Parkinson’s Disease (PD) for less than 5 years (<5y), or 5 years and above (≥5y), and healthy control (HC) subjects. Changes in cell type frequencies (arrows in the cell ratio columns) and surface receptor expressions (drawings on each cell type ⸸) are shown for the innate immune cells in the PBMC pool: for the overall population of TLR2+ (inclusion marker) mononuclear phagocytes (MNPs) and uniquely for each subtype of monocytes (classical (cMos), intermediate (iMos), and non-classical (ncMos)), dendritic cells (DCs), and (bottom right frame) the TLR2-/CD56dim mature natural killer (NK) cells, which had an increase of CD16 expression in PD with a timing difference for the sexes. Sex-differences (far right column) were also observed for the expression levels of TLR4, CCR2, and CD11b with arrows indicating the direction of the change when comparing (early) PD vs. HC. The inclusion marker for MNPs, TLR2 expression, did not change according to disease duration, but to Hohn and Yahr stage (H&Y). ⸸ For simplicity, the depicturing of CD163 on MNPs and CD11b on cMos refers to percentages of cells positive for these receptors and not the expression of these receptors per cell (MFI). Illustration was made using BioRender.com.
Interestingly, fewer CD11b-/CCR2dim and ncMos, and more CCR2bright MNPs and cMos (all in late PD), and increased CCR2 expression (in females), as well as higher MNP viability were all related to worse cognition in PwP. Similarly, when analyzing according to cognitive MoCA scores, both the expression level of CCR2 (females only) and CD11b on iMos were also higher in patients with cognitive impairment. Furthermore, we showed an upregulation of the α-syn sensor, TLR2, in patients with a H&Y score above 2. This was reflected in increased TLR2 expression in follow up visits in the longitudinal cases. Altogether, we showed peripheral innate immune changes during early PD with increases of cells and proteins related to immune activation, phagocytosis, and extravasation and correlated with symptoms. While some of these changes were maintained when the disease duration extended beyond 5 years, late PD, many faded maybe as a sign of exhaustion. Some changes were sex-dependent, suggesting sex-differential immune response during the PD neuroimmune process. Therefore, our study warrants future sex-stratified longitudinal studies.
The distribution of Mo subtypes has previously been investigated with varying results. Some reported no changes (Alvarez-Luquin et al., 2019; da Silva et al., 2016; Drouin-Ouellet et al., 2014; Schlachetzki et al., 2018; Schroder et al., 2018). One of these studies included iMos in the ncMos gate (all CD16+), which might explain their increased “ncMos” in PD CSF (Schroder et al., 2018). In contrast, another study, including iMos in the cMos gate (all CD14+), reported increased cMos and decreased ncMos in PwP vs. controls (Grozdanov et al., 2014). Moreover, a recent study on 41 early PD cases described increased cMos, decreased iMos and ncMos driven by patients with a high risk of dementia (Wijeyekoon et al., 2020b). Notably, none of these studies analyzed the DCs. The lack of consensus might be related to differences in the stages and types of PD, medication, comorbidities, sex, and flow gating strategy (Marquez et al., 2020; Umehara et al., 2020). Here, as a unique approach, we pre-gated MNPs using TLR2. The sporadic PD cases included were L-DOPA treated (with few exceptions, Table 1), sex-balanced, and with similar prevalence of anti-inflammatory treatment as HCs (Supp.Table 2). This approach and the early/late division revealed changes related to PD stages, i.e. disease duration, with a pattern of contraction of the cMos in late vs. early PD with a parallel increase on ncMos (2-way ANOVA). The apparent early cMos expansion is in line with our previous observation of increased cMos in patients with idiopathic REM sleep behavioral disorder (iRBD), considered prodromal PD (Farmen et al., 2021). We observed, however, a late stage increase of iMos, agreeing with a recent work showing increase iMos in PwP. Although in their cohort it was mostly driven by patients at H&Y stage 4, and without information on disease duration, making it difficult to compare with our study directly (Thome et al., 2021). We observed significant decrease in DCs, corroborating prior reports of reduced DCs in PwP vs. HCs (Alvarez-Luquin et al., 2019; Ciaramella et al., 2013; Goldeck et al., 2016).
Previously, we observed increased CD56dim mature NK cells in iRBD patients (Farmen et al., 2021), and others reported increased NK cells, and NK activity in PwP (Mihara et al., 2008; Niwa et al., 2012). Here, PwP had mature NK cells with increased activation as revealed by higher CD16 expression that occurs early in females and late in males PwP. This might suggest an enhanced antibody-dependent cellular cytotoxicity with a differential temporal evolution in each sex. Interestingly, IgG deposition on neurons in close proximity to CD16+ (FcγRIII) lymphocytic cells has been shown in post-mortem PD brains (Orr et al., 2005). Hence, the increased CD16 expression on NK cells might contribute to adaptive immunity-mediated neurodegeneration. However, future functional analyses are needed to evaluate this hypothesis.
Further suggesting a role for adaptive immunity, we observed increased expression in HLA-DR, which presents antigen-peptides to Th-cells, thus linking innate and adaptive immune components. Increased HLA-DR/MHC-II expression was found on Mos in the CSF from PwP (Fiszer et al., 1994), in post-mortem PD brains (Imamura et al., 2003b; McGeer et al., 1988), and PD models (Barkholt et al., 2012; Harms et al., 2013; Thomsen et al., 2021). Correspondingly, we found HLA-DR expression increased on iMos and ncMos in late PD not in early; in agreement with two recent papers showing similar HLA-DR increase (Thome et al., 2021), and one reporting no changes in early PD (Wijeyekoon et al., 2020b). Moreover, we previously observed decreased HLA-DR expression on RBD patients’ MNPs (Farmen et al., 2021) suggesting that HLA-DR expression might be differentially modulated in each disease stage (prodromal, early, and late). A note of caution: the increased HLA-DR expression was also associated with L-DOPA doses, which increase with disease duration and as a consequence of the disease progression in PwP. Thus, it is unclear whether this HLA-DR upregulation is indicative of a PD-related increased antigen presentation capacity, or a molecular event induced by the L-DOPA. This will require further investigation.
We and others have shown that MNP-related changes seem particularly relevant for cognition in PD (Ciaramella et al., 2013; Nissen et al., 2021; Wijeyekoon et al., 2020b). Elevated CCR2 expression is a signature of immature Mos marked for migration to blood/tissue. Increased Mo-precursor cells/cMos expressing CCR2 have been described in PD (Funk et al., 2013; Patel et al., 2017) and increased leukocyte CCR2 mRNA expression is associated with cognitive decline (Harries et al., 2012). Here, we showed increased CD11b (in males) and CCR2 expression (in females) and increased CCR2+/CD11b+ and CCR2+/CD163+ double-positive MNPs in early PD. Our complex analysis informed about MNPs co-expressing multiple markers contributing to PD, rather than CCR2 alone. Cohort#1A included mostly PwP with no (MoCA>26) or (MoCA=18-26) mild cognitive impairment, only three had MoCA<18 = (mild) dementia. Despite these relatively high MoCA scores, we observed that the combined absence of both migration markers was associated with better cognition, and accordingly, increased CCR2+ MNP percentage (in late PD) or higher CCR2 expression (in females) correlated with impaired cognition. Similarly, for the late patients we observed that an imbalance of Mo subtypes (increased cMos and reduced ncMos) was associated with worse cognition. Furthermore, the iMos’ MFI of CD11b and CCR2 (females only) was higher in those patients with a MoCA scores indicative of cognitive impairment (MoCA ≤26). This corroborates that Mos with migratory capacity are associated with cognitive decline in PwP (Nissen et al., 2021; Umehara et al., 2020; Wijeyekoon et al., 2020b). Therefore, indicating that a long-term Mo dysregulation is disadvantageous for the patients, which deserves further investigation in a cohort with severe cognitive impairment.
Immunological sexual-dimorphism, including sex-divergent Mo aging (Marquez et al., 2020), has been described (Bouman et al., 2004; Sue, 2017). Some sex differences seem constitutive, such as the higher female TLR4 expression observed here in HCs. Whereas, sexual-dimorphism for CCR2 and CD11b were obvious only in PwP. The higher TLR4 expression in female HCs might be particularly relevant due to TLR4’s protective role by clearing α-syn (Choi et al., 2020; Earls and Lee, 2020), which could contribute to the lower PD risk for females. However, prolonged misfolded α-syn-sensing by TLR4 also contributes to (chronic) inflammation (Campolo et al., 2019), which might be differently regulated in men and women. The relevance of TLR4 is reinforced by our previous data showing Mo TLR4 expression positively correlating with midbrain immune activation and decreased dopaminergic putaminal transmission in iRBD (male) patients (Farmen et al., 2021). In that iRBD cohort, TLR2 expression was correlated with hyposmia. Moreover, here in our cross-sectional Cohort#1A, we detected a significant increased TLR2 expression on iMos, ncMos, and DCs in patients with H&Y score above 2 vs. those with lower scores. Likewise, we observed increasing TLR2 expression on MNPs, cMos, and ncMos in our longitudinal case reports (Cohort#1B). Although previously reported data on TLRs in other PD cohorts are conflicting (da Silva et al., 2016; Drouin-Ouellet et al., 2014; Wijeyekoon et al., 2020b), our prior data in iRBD (Farmen et al., 2021) and the observations here in PwP confirm the relevant role of TLR2/4 in PD-pathology suggested previously by human, animal and cellular studies (Harms et al., 2021).
TLR activation and other inflammatory events induce shedding of the MNP-specific membrane receptor CD163, forming sCD163 (Moller, 2012). Plasma sCD163 correlates negatively with CD163 MFI, but not with CD163+ cell counts (Davis and Zarev, 2005), thus, increased sCD163 paralleled a decreased CD163 MFI in multiple sclerosis (Gjelstrup et al., 2018). With our previous finding of increased serum sCD163 levels in females with PD from another cohort (Nissen et al., 2021), we anticipated decreased CD163 MFI on MNPs from females with PD in the Cohort#1A (same biobank, different donors). However, we observed significantly increased CD163 MFI in PwP for both sexes, indicating a PD-associated upregulation of CD163 that would parallel the increased shedding in serum (females) and CSF (both sexes) that we described before (Nissen et al., 2021). Although CD163 upregulation was not observed in a cohort of iRBD (male) patients, increased CD163+ cell frequency was associated with reduced immune activation in the midbrain and preserved dopaminergic neurotransmission in putamen both shown by PET imaging (Farmen et al., 2021). This was suggestive of a protective role of the CD163 population at this prodromal PD stage. Indeed, CD163 expression has previously been associated with alternative activation of MNPs considered of anti-inflammatory character (Fischer-Riepe et al., 2020).
The PD-related CD163 upregulation and co-expression with CD11b and CCR2 suggests CD163+ cell infiltration into CNS. Infiltrated CD163+ cells are found in the post-mortem brains of PD models and PwP (Harms et al., 2017; Pey et al., 2014a; Tentillier et al., 2016). The brain infiltration and subsequent activation of the CD163+ cells may lead to sCD163 production, in agreement with our previously reported increased sCD163 in CSF from PwP (Nissen et al., 2021). Furthermore, the CSF-sCD163 levels in PwP were correlated with markers of angiogenesis and chemotaxis, such as ICAM-1 (CD11b agonist) and CCL2 (CCR2 ligand)), CSF-α-syn, and cognition (Nissen et al., 2021). Altogether, our current and prior data suggest that CD163+/CCR2+/CD11b+ cells seem to infiltrate the brain of PwP, differentiate into MΦs, and influence the CNS inflammatory status and neurodegeneration. However, the CD163 receptor’s specific role in PD remains to be determined.
The fewer HCs might limit the present study. Additionally, alternative separation based on PD subtypes or progression rates (Wijeyekoon et al., 2020b) might challenge our early/late-division that is based on disease duration. Nevertheless, this division revealed otherwise hidden differences, which could explain previous inter-study MNP inconsistencies in PD. Moreover, this did not preclude us to associate changes with scores related to disease severity (MoCA and H&Y).
5. Conclusion
Altogether, our study suggests that the PD immune response differs with disease duration/stages by activation in early PD and some degree of immune-exhaustion or stabilization emerging at late-stage. We show a longitudinal increase of TLR2 on MNPs in PwP, related to motor worsening by H&Y in the cross-sectional cohort. Our data indicate stage (disease duration)-associated alterations in the innate immune compartment, particularly in the phagocytic CD163+ migration competent CCR2+/CD11b+ cMos and iMos, but for all TLR2+ MNPs in general. We observed increases of innate immune cells and receptors in especially early PD, while HLA-DR was upregulated mainly in late PD. We also showed significant increased CD16 expression on mature NK cells, relevant for antibody-dependent cytotoxicity, with a different temporal evolution in male and females. Further highlighting the sex dimorphism observed in PD, we also observed sex related TLR4, CCR2, and CD11b differences. Interestingly, many of these changes were related to symptoms of PD: the associations of TLR2 with the motor symptoms, DCs with hyposmia, plus CCR2 and CD11b with cognition. Altogether, our findings show alterations in the peripheral immune system in PD related to the disease duration and highlight a need to further study CD163+ MNPs, focusing on the role of cMos and iMos in PD.
Data Availability
The data supporting the conclusions are included within the article and its additional file.
Data Availability
The data supporting the conclusions are included within the article and its additional file.
6. Declarations
6.1 Ethics approval and consent to participate
The study was approved by the Ethics Committee of the University of Tuebingen (Germany) committee (480/2015BO2). All participants providing written informed consent.
6.2 Availability of data and materials
The data supporting the conclusions are included within the article and its additional file.
6.3 Financial disclosure during the past 12 months
K. Brockmann has received a research grant from the University of Tuebingen (Clinician Scientist), the dPV, the MJFF, and the German Centre for Neurodegenerative Diseases (DZNE, MIGAP); travel grants from the Movement Disorders Society, and speaker honoraria from Abbvie, Lundbeck, UCB, and Zambon.
M. Romero-Ramos serves on the editorial board of Brain Research and npj Parkinson’ disease, and receives research support from Novo Nordisk Foundation, Aarhus University Research Foundation, the Danish Medical Research Council, the Danish Parkinson Foundation, Desiree and Niels Ydes Foundation, and the Michael J Fox Foundation.
SKN, KF, MC, CS, and DG report no disclosures.
6.4 Authors’ contribution
SKN: Study design, flow of Cohort#1A, data analysis, wrote the first draft ; KF: Flow and analysis of Cohort#1B; MC: Flow; CS: Coordinated sample selection and collected biobank archive info; DG: isolated PBMCs; KB: Supervised the biobank; MRR: Developed the concept and designed the study. MRR & SKN: Interpreted the data and wrote jointly the final version. All authors critically revised the manuscript.
Financial disclosure
The authors declare no conflict of interest.
Funding sources
Funding support for the research covered by this article was provided by the Michael J. Fox Foundation (MRR), the Bjarne Saxhof Fund administered through the Danish Parkinson’s Foundation (MRR) and the Danish Council for Independent Research (MRR). The authors declare that they have no competing interests.
6.5 Acknowledgments
We acknowledge the invaluable technical help provided by Gitte Ulbjerg Toft (Department of Biomedicine, Aarhus University). Samples were obtained from the Neuro-Biobank of the University of Tuebingen, Germany (https://www.hih-tuebingen.de/en/about-us/core-facilities/biobank/ ), which is supported by the local University, the Hertie Institute and the DZNE. Flow cytometry was performed at the FACS Core Facility, Aarhus University, Denmark.
Footnotes
From the original analyzed samples (n=109), three individuals with known autoimmune comorbidities (AID) were excluded (1 female control and 2 late PD, one female and one male) in this version of the manuscript. This changed the statistic for some findings. We applied additional analysis of Hohn and Yahr scoring with 2 as a cutoff, and MoCA scoring with 26 as a cutoff, revealing an association between high expression of immune receptors and worse PD symptoms.
7. References




