
Abstract
Objectives The gut microbiome modulates human brain function and eating behavior through multiple factors, including short-chain fatty acid (SCFA) signaling. We aimed to determine which bacterial genera relate to eating behavior, diet and SCFA metabolites in overweight adults. In addition, we tested whether eating-related microbiota predict treatment success in patients after bariatric surgery.
Design Anthropometrics, eating behaviour measures and 16S-rRNA-derived microbiota abundance were correlated in young overweight adults (n=27 (9F), 21-36 years, BMI 25-31 kg/m2), and in patients two years after Roux-en-Y gastric bypass (n=23 (16F), 41-70 years, BMI 25-62 kg/m2) compared to matched overweight and obese controls (n=17 (11F), 26-69 years, BMI 25-48 kg/m2). Dietary fiber intake was assessed and SCFA were measured in feces and blood using mass spectrometry in young adults.
Results In the young adults, 7 bacteria genera, i.e. Alistipes, Blautia, Clostridiales cluster XVIII, Gemmiger, Roseburia, Ruminococcus and Streptococcus, correlated with healthier eating behavior, while 5 genera, i.e. Clostridiales cluster IV and XIVb, Collinsella, Fusicatenibacter and Parabacteroides, correlated with unhealthier eating (all |r| > 0.4, FDR-corrected p < 0.05). Some of these genera including Parabacteroides related to fiber intake and SCFA metabolites, and to weight status and treatment response in obese patients.
Conclusion Specific bacterial genera, particularly Parabacteroides, were reliably associated with weight status and eating behavior in two independent well-characterized cross-sectional samples. Together, these findings indicate two groups of presumably beneficial and unfavourable genera that relate to eating behaviour and weight status, and indicate that dietary fiber and SCFA metabolism may modify these relationships.
Significance of this study What is already known about this subject?
- The gut microbiome may modulate brain function and eating behavior through production of short-chain fatty acids (SCFA) from dietary fiber.
What are the new findings?
- This cross-sectional exploratory study finds that the abundance of specific groups of bacterial genera, including Parabacteroides, correlate with eating behavior, diet, SCFA as well as weight status and obesity treatment success in two independent well-characterized samples.
How might it impact on clinical practice in the foreseeable future?
- These findings might help to develop novel hypotheses how to prevent and treat unhealthy food craving through microbiotal modulation of the gut-brain axis.
Introduction
Gut microbes modulate brain function and behavior via immune, endocrine, neural and humoral routes [1]. This could play a key role in neuronal feeding circuits and overeating, as dysbiosis of the microbiota composition has been documented in psychiatric eating disorders [2] and obesity [3].
However, nutrition-or body weight-related microbial changes and their functional relevance are still relatively unclear. In mice, gastric bypass-related differences in the microbiota profile, such as a higher abundance of the genera Escherichia (phylum Proteobacteria) and Akkermansia (phylum Verrucomicrobia), induced weight loss when transferred to germ-free animals [4]. In humans, bariatric surgery similarly led to higher overall microbiota diversity and to higher abundance of the species Escherichia coli and in some studies to further abundance changes within the phylum Bacteroidetes, such as a higher post-surgery ratio of the genera Bacteroides to Prevotella [5] and less Firmicutes (phylum level) or to more Gammaproteobacteria (class level) [6]. The ratio of Bacteroides to Prevotella at baseline predicted dietary weight loss success after 24 weeks in an intervention study in 80 overweight individuals [7]. Further, a one-week dietary intervention trial in 20 individuals found that microbial composition predicted glycemic response [8].
Human-to-mouse fecal transplant experiments further underline the causal role of specific microbiota to facilitate weight loss [9], and human-to-human microbiota transplantation experiments increased insulin sensitivity according to [10]. In a recent human study, accompanied by mouse model data, an individual’s microbiota profile, extracted from fecal samples during periods of dietary weight loss, prevented weight regain when transferred back to the individuum orally [11].
Mechanistic insights into how specific gut bacteria modulate human eating behaviour and weight status are still limited. The gut microbiota is supposed to affect the host’s metabolism by altering energy extraction from food, and by modulating dietary or host-derived compounds that modify the metabolic pathways of the host [12]. For example, short-chain fatty acids (SCFA) are excreted by certain gut bacteria as a result of carbohydrate fermentation, and SCFA stimulate the secretion of anorexigenic hormones, such as peptide YY (peptide tyrosine tyrosine or PYY) and glucagon-like-peptide-1 (GLP-1) in the colon, which further signal to hypothalamic nuclei as one mechanism of homeostatic regulation [13]. SCFA can also cross the blood-brain-barrier and act as signaling molecules in the brain to directly modulate appetite and food-decision making [1]. First interventional studies showed that intake of butyrate (one type of SCFA) or the butyrate-producing bacteria Akkermansia spp. exert beneficial effects on body weight depending on treatment intention in humans [14] and on brain functions in mice [15], including reduced food intake [16]. Notably, specific pre-biotic nutrients, such as dietary fibers, are known to nourish SCFA-producing bacteria in the gut, rendering diet a potent modifier of gut-brain signalling [17]. In sum, the gut microbiome may influence feeding behaviour, e.g. by modulating reward and homeostatic signaling [18,19] and by stimulating the vagal nerve [20], in particular in dysregulated biological systems, such as in food addiction [21] or eating disorders [2]. Yet, direct knowledge if specific genera are linked to eating behavior via dietary intake and SCFA in humans is lacking. Here, we asked whether gut microbial diversity and genera abundance relate to eating behaviour, and to SCFA metabolites in the colon (feces) and in the periphery (blood) in a homogenous sample of young overweight adults. In addition, we tested whether the abundance of eating behavior-related microbiota differed between overweight and obesity in a sample of older adults and if they predict treatment success (i.e. achieved weight loss) in obese patients at two years after bariatric surgery.
Methods
Samples characteristics and data collection
We included cross-sectional data from two longitudinal studies, i.e. 1) healthy young overweight adults (n = 27 (9F), 21-36 years, BMI 25-31 kg/m2) (Clinical Trials registration NCT03829189), and 2) patients two years after Roux-en-Y gastric bypass (RYGB) surgery, with “good” (n = 11 (7F), 41-70 years, BMI 25-29 kg/m2) and “bad”, i.e. only little weight loss response (n = 12 (9F), 31-67 years, BMI 41-62 kg/m2), as well as age-, gender-and BMI-matched controls (overweight: n = 8 (5F), 41-58 years, BMI 25-29 kg/m2; obese: n = 9 (6F), 26-70 years, BMI 41-48 kg/m2). All participants completed anthropometric measurements and donated feces (see SI) for microbiota analysis (Shannon effective [22] and relative abundance of microbiota genera), underwent anthropometric measurements and filled in questionnaires to quantify eating behaviour traits. Sample 1 additionally provided blood, and SCFA (acetate, butyrate, propionate) were measured in feces and serum. Also, for sample 1, self-reported dietary fiber intake (g/day), computed from a standardized food frequency questionnaire and hunger ratings after overnight fasting and post-prandial to a standardized meal (10% of caloric need) (methods similar to [23]) were assessed.
Microbiota assessment
To assess microbiota community structure we used 16S rRNA gene profiling of the fecal samples. Therefore, DNA was extracted and V3-V4 variable regions of the 16S rRNA genes were amplified by PCR and a library was constructed, followed by paired end 2×250bp Illumina sequencing. These analyses were done by GENEWIZ Germany GmbH, Leipzig). Next, the inhouse Galaxy server using a pipeline implemented with the DADA2 R package processed raw data in fastq format. For each sample, paired-end reads were joined, low-quality reads were removed, reads were corrected, chimeras removed and Amplicon Sequence Variants (ASVs) were obtained. Taxonomy was annotated to the ASVs using the RDP database [24]. The read counts per ASV with taxonomic annotation were normalized and relative abundances of each ASV and taxa were calculated using the R scripts Rhea. Visualization was done as in [25] by inhouse written R-tools using the ggplot2 R-package.
Eating behaviour
To characterise eating behaviour traits, questionnaires based on self-report were used: the Three-Factor Eating Questionnaire (TFE-Q) (German version, [26]) and the Eating Disorder Examination Questionnaire (EDE-Q) (German version, [27]) were used for sample 1 and sample 2, respectively. The TFE-Q assesses three domains of eating behaviour (cognitive restraint, disinhibition, hunger), and the EDE-Q covers the subscales dietary restraint, eating concern, weight concern, and shape concern. Scoring was performed according to the respective manuals.
Additional analyses in sample 1
Hunger ratings and health indicators, i.e. blood pressure and body fat, as well as dietary intake were assessed (see SI). Blood was obtained in fasting state (12 ± 3h fasted) and samples were centrifuged at 3500 revolutions per minute at 7°C for 6 minutes. Serum was aliquoted within one hour of obtainment. Processed aliquots were stored at −80°C until data analysis. SCFA in blood and stool were analysed according to [28] (see SI).
Obesity surgery in sample 2
For sample 2, 23 morbidly obese patients were drawn from a larger pool of RYGB (see SI) patients from the University Hospital Leipzig, Germany (ethics proposal 027/17-ek), based on their response to the surgical treatment. Specifically, patients were identified from the database if their surgery dated back at least 2 years (mean 53 months ± 16.5 SD, range 25-76 months) and those were further divided in percentiles according to relative excessive weight loss (EWL) (good responders: sustained EWL > 60 %, mean 93% ± 4 SD, range 86-98%, n = 12; bad responders: sustained EWL < 40 %, mean 20% ± 13 SD, range 3-35%, n = 11).
Statistical analysis
Correlational analysis
Relative taxa abundance (%) on the genera level was used as primary variables of interest. All analyses were performed in RStudio version 3.6.1. Non-normally distributed variables were log-or Tukey-transformed. For the main analysis, 20 out of 120 genera were included as they appeared in at least 80% of individuals [29] and fed into a correlation matrix with all main variables of interest in sample 1, i.e. TFEQ traits, hunger ratings, body fat, systolic blood pressure, dietary fiber intake and SCFA in feces and blood. All values were FDR-corrected and the statistical significance was set to p < 0.05. Those genera that were significantly associated with eating behavior (TFEQ traits and hunger ratings, FDR-corrected, p < 0.05) were then correlated with weight status and RYGB treatment success in sample 2. Group differences across overweight, obese, good and bad RYGB responders were tested with non-parametric Kruskal-Wallis tests. Further correlations were tested with Pearson’s correlation coefficient r for normally distributed variables or with Spearman’s rho for non-normally distributed variables.
To further investigate, if the interplay of correlated genera -rather as a holobiont than individually -is determinative of the observed relations, the relation between correlated to non-correlated genera was computed by three composite scores (1)-(3).
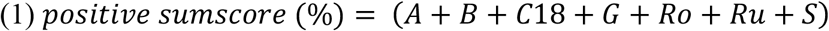
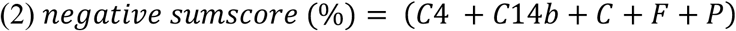
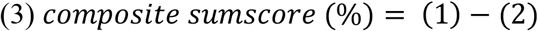 Here, higher scores signify an overall healthier profile with regard to eating behaviour.
Here, higher scores signify an overall healthier profile with regard to eating behaviour.
Mediation Analysis
Using simple mediation analysis with in RStudio version 3.6.1 using medmod() we checked for statistical mediation in sample 1 for variables showing bivariate correlations in the following paths:
fiber —> correlated genera or sumscores —> eating behaviour (TFEQ, hunger ratings)
eating behaviour (TFEQ, hunger ratings) —> fiber —> correlated genera or sumscores
correlated genera or sumscores —> SCFA —> eating behaviour (TFEQ, hunger ratings)
Significance was set to p < 0.05, and in the main analysis for sample 1 we corrected for multiple testing using the false-detection rate (FDR)-correction.
Results
Characteristics of sample 1 and 2 are listed below (see Table 1-2). Eating behaviour traits varied across both samples, and in sample 2, restrained eating and shape/weight concerns differed between those that achieved long-term excessive weight loss after bariatric surgery compared to those that did not (good vs. bad responders, all W> 58.5, p < 0.001, Tab. 2, Suppl. Figures 1).
Descriptives for sample 1.
Descriptives for sample 2.
Overall microbiota diversity at the phylum level was relatively comparable across participants of samples 1 and 2 except higher ratio of Firmicutes to Bacteroidetes in sample 1, and Prevotellaceae and Fusobacteriaceae families were more abundant in patients after RYGB surgery (Figure 1, see SI for details).

Microbiota, eating behaviour traits and health indicators in overweight adults
In sample 1, higher relative abundance of Collinsella (phylum Actinobacteria), Clostridium IV and XIVb, Fusicatenibacter (all three phylum Firmicutes) and Parabacteroides (phylum Bacteroidetes) were related to less healthy eating behaviour (higher TFEQ scores and/or higher hunger ratings, all 0.61 < |r| > 0.42, p-FDR < 0.05, Figure 2A). Contrastingly, higher relative abundance of the microbial genera Alistipes (phylum Bacteroidetes), Blautia, Clostridium XVIII, Gemmiger, Roseburia, Ruminococcus and Streptococcus (all phylum Firmicutes) correlated with healthier eating behaviour (all 0.76 < |r| > 0.42, p-FDR < 0.05, Figure 2B, Suppl. Figure 4).

Further, Collinsella abundance significantly correlated with higher body fat mass (sex-standardized, r = 0.61, p < 0.001, Figure 2C). Streptococcus abundance was significantly correlated with lower mean systolic blood pressure (r = −0.70, p-FDR < 0.001, Figure 2D).
Relation to dietary fiber intake and SCFA
Out of the 12 genera that were significantly associated with eating behaviour (from now on called “(inversely) health-related” genera), three were associated with lower (Collinsella and Parabacteroides) or higher (Clostridium XVIII) dietary fiber intake (all 0.73 < |r| > 0.49, p-FDR < 0.05, Figure 3A-C). Moreover, higher dietary fiber intake per se was significantly associated with lower disinhibited eating (r = −0.58, p-FDR < 0.01) and lower body fat mass (r = −0.75, p-FDR < 0.0001, Figure 3D-E).

SCFA concentrations in feces were highly variable and up to ∼1000 times higher in serum for all three measured SCFA (all t(24) > 11.6, p < 0.001). Serum acetate was 2.5 times higher compared to butyrate and propionate in serum (Suppl. Table 1).
We observed that higher abundance of some of the inversely health-related genera correlated with higher levels of different SCFA in feces and serum (all r > 0.50, p-FDR < 0.01). In addition, most health-related genera correlated with some feces and serum SCFA markers, however revealing both positive and negative associations (only those associated with eating behaviour were considered, all 0.65 < |r| > 0.44, p-FDR < 0.05, Suppl. Figure 5). Note, that some genera showed differential correlations within the different SCFA, e.g. higher Alistipes correlated with higher acetate in both feces and serum and with fecal butyrate, but with lower fecal propionate. Moreover, considering the inversely correlated genera, Fusicatenibacter and Parabacteroides correlated significantly with higher fecal concentrations of propionate and acetate, respectively.
Also, higher fecal propionate levels correlated significantly with higher cognitive restraint eating (r = 0.50, p-FDR = 0.014, Figure 4A). Higher fecal acetate, butyrate and propionate levels correlated with higher hunger ratings (all r > 0.45, all p-FDR < 0.04), but also serum propionate with hunger (r = 0.45, p-FDR = 0.03). Moreover, serum acetate and butyrate were inversely associated with body fat mass (all r > −0.43, all p-FDR < 0.04) (Figure 4B-C). Notably, serum levels did not correlate with fecal SCFA concentrations (all r < | 0.17 |, all p-uncorr < 0.86, Suppl. Figure 6).

Genera sum score and mediation analyses
The negative sumscore of the five inversely health-related genera abundance resulted in significant correlations for two of the eating traits (cognitive restraint r = 0.59, p-uncorr = 0.001; disinhibition r = 0.65, p-uncorr < 0.001, Figure 5A-B). The positive sumscore of the seven health-related genera showed no significant associations (all p-uncorr < 0.95, Suppl. Figure 7). Neither sumscore correlated with fecal or serum SCFA levels.

Exploratory mediation path analyses of the proposed models did not show statistically significant mediating paths for differences in diet, eating behaviour or hunger ratings through differences in Parabacteroides or positive/negative sumscores (Suppl. Table 2-3). Considering SCFA, similar results emerged, except for acetate: Here, while the direct effect c’ did not reach significance (β = −0.3, p = 0.13), higher Parabacteroides abundance was linked with higher postprandial hunger ratings through higher fecal acetate levels (indirect effect, a*b, β = 0.36, 95% CI [0.05 0.66], p = 0.02, Suppl. Table 4).
Mircobiota genera differences between overweight, obese and surgery groups
In sample 2, two of the five inversely health-related genera identified in sample 1 were significantly different between groups (all H(3) > 9.5, p < 0.023) with lower relative abundance of Parabacteroides in good vs. bad responders (H(1) = 4.9, p = 0.027) Figure 6). In addition, six of the seven health-related genera identified in sample 1 were more abundant in the overweight group (all H(3) > 8.3, p < 0.036, Figure 7), but did not differ in the good vs. bad RYGB responders.


Considering the sumscores, we found that both sumscores differed between groups (Figure 8, all H(3) > 11.3, p < 0.01) with the negative sumscore showing higher values in the bad vs. good RYGB responders (H(1) = 2.1, p = 0.036). In addition, both the positive (n.s.; H(1) = 1.9, p = 0.05) and the negative sumscore (H(1) = 2.02, p = 0.043) showed higher values in overweight vs. obese participants.

Bad vs. good RYGB responders showed higher eating restraint scores (H = 5.3, p = 0.022, Suppl. Figure 1), and higher scores correlated with higher Parabacteroides abundance in these groups (r = 0.44, p = 0.039, Figure 9A). Moreover, lower Parabacteroides abundance correlated significantly with higher weight loss after surgery (r = 0.49, p = 0.019, n = 20, Figure 9B). The negative sumscore correlated with unhealthier eating behaviour (Figure 9C, mean of all EDEQ subscales, r = 0.47, p = 0.027; EDEQ restraint, r = 0.49, p = 0.022) and with less weight loss after surgery (Figure 9D, weight, r = 0.53, p = 0.011, BMI, r = 0.53, p = 0.011).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
RYGB, Roux-en-Y gastric bypass.
Discussion
Combining data from two human cross-sectional datasets, we find two groups of microbiota genera that were either positively or inversely associated with both healthier eating behaviour and anthropometrics (1) in a deeply phenotyped sample of young overweight adults and (2) when comparing patients showing a good or bad response two years after bariatric surgery with matched controls, respectively. More specifically, in young overweight adults, 7 bacterial genera, i.e. Alistipes, Blautia, Clostridium XVIII, Gemmiger, Roseburia, Ruminococcus and Streptococcus, correlated with healthier eating behaviour traits and lower subjective hunger ratings, indicating potential benefits for the host metabolism, while 5 bacterial genera, i.e. Clostridum IV, Clostridium XIVb, Collinsella, Fusicatenibacter and Parabacteroides, correlated with unhealthier eating traits and higher subjective hunger ratings. Collinsella was further related to higher body fat mass and Streptococcus to lower systolic blood pressure. The positively health-related bacterial genera were also more abundant in the overweight good responder controls, compared to the obese bad responder controls and RYGB-operated patients, while the negatively health-related genera showed a less clear distribution across groups, with Parabacteroides being significantly less abundant in good vs. bad RYGB-operated patients. Moreover, relative abundance of Parabacteroides as well as a composite score of all negatively correlated genera, were associated with higher eating restraint and with lower post-operative weight loss across both RYGB groups. Considering diet and SCFA-related pathways, we observed that higher dietary fiber intake in overweight adults correlated with more abundant Clostridium XVIII, and less abundant Collinsella and Parabacteroides, as well as with healthier eating behaviour and anthropometrics. While SCFA showed a rather mixed pattern of correlations with the different markers, Fusicatenibacter and Parabacteroides abundance correlated with higher fecal propionate and acetate, respectively, that again correlated with elevated hunger. Contrastingly, higher acetate and butyrate in serum correlated with lower fat mass, indicating a possible inverse association of acetate in feces and serum with respect to health indicators. Together, these results indicate that presumably beneficial and unfavourable microbiota genera relate to eating behaviour and weight status, and that dietary fiber intake and SCFA metabolism may modify these relationships.
Bacterial genera
The positively correlated microbiota group is comprised of bacterial genera that have been described as beneficial for the host in previous literature. For example, Alistipes and Blautia were found to produce SCFA [30,31]. Similarily, Gemmiger, Roseburia, and Ruminococcus belong to the families of Ruminococcaceae or Lachnospiraceae, which share a common role as active plant degraders [32]. These positive metabolic effects on the host could eventually contribute to improved adiposity control, as e.g. higher Blautia was correlated to lower body fat [33], Roseburia was linked to lower blood glucose and Ruminococcus to higher weight loss in mice after vertical sleeve gastrectomy via regulation of nuclear receptor binding of bile acids [34]. A microbial transfer study from human to mice showed obesity-promoting effects of the species C. ramosum, which is part of Clostridium XVIII [35]. However, studies on the genera Clostridium XVIII and Streptococcus in relation to health are scarce. Clostridia are known to be key commensals for gut homeostasis [36], but classification of the genus Clostridium remains challenging due to the high heterogeneity of the listed species [37]. Also, there are currently 50 species identified in the genus Streptococcus alone, rendering different functionality in these genera likely. Yet, we found that Clostridium XVIII abundance related to higher dietary fiber intake, and Streptococcus abundance to lower blood pressure. Indeed, fiber intake related to healthy eating behaviour and lower body fat mass in overweight young adults in the present analyses may point towards rather beneficial fiber-correlating Clostridium XVIII and Streptococcus genera species that underly those associations. Moreover, these results underline the potential impact of a fiber-rich diet for health indicators. Due to the exclusive occurrence of fiber in plants, fiber-rich diets are oftentimes attributed to plant-based (vegetarian or vegan) diets, and plant-based diets have been shown extensively to be beneficial for weight status, gut and overall health [38,39].
Considering the negatively correlated group of microbiota, some genera were described to include pathogens, e.g. in Clostridium XIVb the species C. piliforme, the causative agent of Tyzzer’s disease [40] and Parabacteroides as an opportunistic pathogen in infectious diseases [41]. Of note, in the Parabacteroides genera, also beneficial species, e.g. P. distasonis, have been described [42]. The anaerobic Collinsella colonizes mucosal surfaces and has recently been reported to degrade potentially toxic food contaminants found in processed foods [43]. While this could be beneficial for the host, unhealthier eating behaviors (such as intake of processed food) and higher body weight could then likely be related to higher abundances of Collinsella. Likewise, studies showed that Collinsella linked to less dietary fiber intake, which is in line with our results in overweight adults, and higher weight loss in cross-sectional [44] and dietary intervention studies [45]. Fusicatenibacter, including the species F. saccharivorans, are strictly anaerobic sugar fermenters, again linking to unhealthier eating behaviour and obesity [46]. The genus Clostridum IV however has rather been reported as beneficial SCFA producers, e.g. the species Faecalibacterium prausnitzii (F. prausnitzii), which play a noticeable role in intestinal homeostasis [47]. Yet again, those genera comprise many different species and it can also be speculated that some bacteria species or genera underlying the observed correlations could have likely been taxonomically misplaced [40]. Taken together, the negatively correlating microbiota genera seem to consist on the one hand of pathogens, indicative of a rather pro-inflammatory milieu in participants with higher weight status, which is well in line with our findings showing that higher Parabacteroides correlated with unhealthier eating traits and poorer weight loss maintenance in RYGB patients. On the other hand, those negatively correlating genera are comprised of those bacteria that metabolize processed food and sugars, again indicative of higher weight and unhealthy eating behaviour. Future studies now need to integrate microbiota data at the species level and randomized interventional trials are required to eventually understand cause and effect of these eating behaviour -microbiota -diet interactions.
SCFA metabolism
We could not establish reliable links between serum and fecal concentrations of those metabolites. The overall weak relationship could have been expected as for example butyrate is rapidly absorbed by the gut mucosa and reaches blood circulation [48], therefore, fecal levels of butyrate may not directly relate to butyrate-producing bacteria abundance nor to serum levels of butyrate. In addition, biosamples of serum and feces were not collected in a time-locked way, therefore a time difference of hours to days might have blurred potential (anti-) correlations. Indeed, it has been shown, that fecal SCFA levels decrease throughout the day due to metabolization and that overnight-fast duration influenced these results [49].
Still, we found that higher fecal SCFA levels (i.e., acetate, butyrate and propionate) linked to higher subjective hunger ratings and also to higher cognitive restraint (i.e. propionate), whereas lower acetate and butyrate in serum correlated with higher fat mass. Statistical path analyses proposed that higher Parabacteroides abundance link to higher hunger through higher fecal acetate. Bearing in mind that higher fecal SCFA levels may indicate less efficient absorption in the gut, leading to lower SCFA availability in serum [50], these findings are somewhat in line with studies showing reduced appetite and less weight gain after acetate intake [1,51]. Note however, that we did not adjust mediation statistics for multiple testings, rendering false positives likely. In addition, it has been discussed that only a minimal fraction of the colon-derived SCFA directly reaches the brain. Instead, more downstream targets of SCFA signalling might be more important for gut-brain communication, such as SCFA-induced release of GLP-1 and PYY at the gut epithelium, modulation of liver metabolism or indirect signaling via the vagus nerve [1]. Future studies could help to further disentangle the different mechanisms at play by assessing further blood-, tissue-or imaging-based biomarkers of these pathways.
Limitations
Firstly, all analyses are exploratory based on cross-sectional data, therefore no conclusions about causal relationships can be drawn. In addition, both samples are limited by size, especially with regard to the number of variables of interest. Due to these constraints, more elaborate statistical analyses (such as structural equation modelling) could not be performed. A major strength of this study is the inclusion of two independent samples integrating next generation sequencing and SCFA metabolomics with psychological markers in well-characterized adults at risk for future weight gain that yielded similar associations of eating behaviour with gut microbiota at the genera level.
Conclusion
The combination of data from cross-sectional samples of overweight, obese and post-bariatric surgery individuals showed robust and multivariate associations between specific bacterial gut genera, particularly beneficial SCFA-producing genera and presumably unfavourable pathogens or sugar-/processed-food digesting bacteria, with anthropometrics, eating traits and dietary fiber intake. While speculative concerning causality, our results propose key microbiota candidates for diet-gut-brain-behaviour interactions in humans and may help to develop novel hypotheses how to prevent and treat unhealthy food craving through microbiotal modulation of the gut-brain axis. Longitudinal and interventional studies integrating metagenomic approaches and functional pathway analysis are needed to disentangle correlation from causality and to further characterize eating behaviour-relevant microbiota genera at the species level.
Data Availability
All data relevant to the study are included in the article or uploaded as supplementary information
Competing Interests Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We would like to thank all participants and staff of the study team, in particular Laura Hesse, Emmy Töws, Linda Grasser and Mathis Lammert. This work was supported by grants of the German Research Foundation, contract grant number CRC 1052 “Obesity mechanisms”, project number 209933838, subprojects A1 (AV), A5 (AH), AOBJ: 624808 (WF), WI 3342/3-1 (AVW), IFB AdiposityDiseases FKZ: 01EO1501 (AH, WF), by the Else Kröner-Fresenius Foundation (WF), by the German Federal Environmental Foundation (EM) and by the Max Planck Society.
References




