
Abstract
The SARS-CoV-2 lineage B.1.1.7, now designated Variant of Concern 202012/01 (VOC) by Public Health England, originated in the UK in late Summer to early Autumn 2020. We examine epidemiological evidence for this VOC having a transmission advantage from several perspectives. First, whole genome sequence data collected from community-based diagnostic testing provides an indication of changing prevalence of different genetic variants through time. Phylodynamic modelling additionally indicates that genetic diversity of this lineage has changed in a manner consistent with exponential growth. Second, we find that changes in VOC frequency inferred from genetic data correspond closely to changes inferred by S-gene target failures (SGTF) in community-based diagnostic PCR testing. Third, we examine growth trends in SGTF and non-SGTF case numbers at local area level across England, and show that the VOC has higher transmissibility than non-VOC lineages, even if the VOC has a different latent period or generation time. Available SGTF data indicate a shift in the age composition of reported cases, with a larger share of under 20 year olds among reported VOC than non-VOC cases. Fourth, we assess the association of VOC frequency with independent estimates of the overall SARS-CoV-2 reproduction number through time. Finally, we fit a semi-mechanistic model directly to local VOC and non-VOC case incidence to estimate the reproduction numbers over time for each. There is a consensus among all analyses that the VOC has a substantial transmission advantage, with the estimated difference in reproduction numbers between VOC and non-VOC ranging between 0.4 and 0.7, and the ratio of reproduction numbers varying between 1.4 and 1.8. We note that these estimates of transmission advantage apply to a period where high levels of social distancing were in place in England; extrapolation to other transmission contexts therefore requires caution.
Introduction
A novel SARS-CoV-2 lineage, originally termed variant B.1.1.7, is rapidly expanding its geographic range and frequency in England. The lineage was detected in November 2020, and likely originated in September 2020 in the South East region of England. As of 20 December 2020, the regions in England with the largest numbers of confirmed cases of the variant are London, the South East, and the East of England. The variant possesses a large number of non-synonymous substitutions of immunologic significance1. The N501Y replacement on the spike protein has been shown to increase ACE2 binding 2,3 and cell infectivity in animal models4, while the P618H replacement on the spike proteins adjoins the furin-cleavage site5. The variant also possesses a deletion at positions 69 and 70 of the spike protein (Δ69-70) which has been associated with diagnostic test failure for the ThermoFisher TaqPath probe targeting the spike protein6. Whilst other variants with Δ69-70 are also circulating in the UK, the absence of detection of the S gene target in an otherwise positive PCR test increasingly appears to be a highly specific marker for the B.1.1.7 lineage. Surveillance data from national community testing (“Pillar 2”) showed a rapid increase in S-gene target failures (SGTF) in PCR testing for SARS-CoV-2 in November and December 2020, and the B.1.1.7 lineage has now been designated Variant of Concern (VOC) 202012/01 by Public Health England (PHE).
Phylogenetic studies carried out by the UK COVID-19 Genomics Consortium (COG-UK)7 provided the first indication that the VOC has an unusual accumulation of substitutions and was growing at a large rate relative to other circulating lineages. Here we analyse VOC whole genomes collected between October and 5 December 2020 and find that the rate of increase in the frequency of VOC is consistent with a transmission advantage over other circulating lineages in the UK. To substantiate these findings, we investigate time trends in the proportion of PCR tests exhibiting SGTF across the UK on ∼275,000 test results as a biomarker of VOC infection, and examine the relationship between local epidemic growth and the frequency of the VOC. We demonstrate that increasing reproduction numbers (‘R’ values) are associated with increased SGTF frequency among reported cases, our biomarker of VOC infection, and confirm this association through a variety of analytical approaches. Critically, we find evidence that non-pharmaceutical interventions (NPIs) were sufficient to control non-VOC lineages to reproduction numbers below 1 during the November 2020 lockdown in England, but that at the same time the NPIs were insufficient to control the VOC.
Origins and expansion of VOC 202012/01
We examined the time and location of sampling of 1,904 VOC whole genomes collected between October and 5 December 2020, combined with a genetic background of 48,128 genomes collected over the same period. Sequences of the VOC were widely distributed across 199 lower tier local authorities (LTLAs) in England, but highly concentrated in the South East (n=875), London (n=636) and East of England (n=293). Relative to this genetic background, the growth of the VOC lineage is consistent with it having a selective advantage over circulating SARS-CoV-2 variants in England (Figure 1A). While rapid growth of the variant was first observed in the South East, similar growth patterns are observed later in London, East of England, and now more generally across England. Across these regions, we estimate similar growth differences between the VOC and non-VOC lineages of +49% to 53% per generation (Supporting Table S1) by fitting a logistic growth model to the frequency of VOC sequence samples through time and adjusting for an approximate mean generation time of SARS-CoV-2 of 6.5 days (see Supporting Methods) 8,9.

Expansion and growth of the VOC 202012/01 lineage. A) The number of UK LTLAs reporting at least one sampled VOC genome. B) Empirical (solid) and estimated (dash) frequency of TPR-adjusted SGTF in three regions of England. C) Empirical (points) and estimated (line) frequency (log odds) of VOC inferred from genomic data by epidemiological week. D) Empirical (points) and estimated (line) frequency (log odds) of SGTF based on the same data as B.
S gene target failure in SARS-CoV-2 testing as a biomarker for the VOC
The UK has a high throughput national testing system for community cases, based in a small number of large laboratories. We were able to extend our genomic analyses to epidemiologic case data, because the VOC lineage is not detected in the S-gene target in an otherwise positive PCR test (ThermoFisher TaqPath as performed in the UK national testing system). Several SARS-CoV-2 variants can result in SGTF, but since mid-November, more than 97% of Pillar 2 PCR tests showing SGTF are due to the VOC lineage10. Before mid-November 2020, the frequency of SGTF among PCR positives was a poorer proxy for frequency of the VOC. We therefore developed a Gaussian Markov Random Field model (see Supplementary Information, Figure S1) to predict the proportion of SGTF cases attributable to the VOC lineage by area and week, here termed the true positive rate (TPR), and the number of SGTF cases attributable to the VOC. In turn, the corresponding false-positives were attributed to the S-gene positive case (S+) category.
Trends in SARS-CoV-2 cases with S gene target failure that are attributed to the VOC
SGTF data were available for 35% of Pillar 2 positive test results between November 26 to December 13, 2020. Given the greater abundance of SGTF data, a more detailed picture of the VOC frequency over time can be discerned after our TPR adjustments. Overall, empirical and estimated frequencies of TPR-adjusted SGTF cases show a similar pattern of expansion as frequencies estimated from genetic data in terms of time, region, and rate of growth (Figure 1D). As of December 13, SGTF is detected in all regions of England (Figure S2), and the estimated frequency of TPR-adjusted SGTF ranges from 15% in Yorkshire and the Humber to 85% in the South East, where the VOC was first detected. Changes in COVID-19 infections correlate with raw (not adjusted for TPR) SGTF cases on a regional basis. Figures 2 and S3 shows the time trends of SGTF (S-) cases, S-gene positive cases (S+) and total PCR positive cases by NHS England Sustainability and Transformation Plan (STP) areas (a geographic subdivision of NHS Regions). Visually, it is clear that while lockdown successfully controlled S+ cases in virtually every STP, S-case numbers increased during lockdown.

Case trends in a subset of NHS STP areas. Total cases reported are shown as a thick line. A subset of these - those tested in the 3 largest “Lighthouse” laboratories - were tested for SGTF. The total cases line is coloured according to percentage S-among those tested. Counts of S+ and S-reported via the PHE SGSS system are shown by the thin lines. The dates of the second lockdown are indicated by the vertical red lines. Nine representative NHS STP areas from all regions of England are ordered by decreasing percentage S-in the most recent week of data. Raw SGTF data are shown here (not adjusted for TPR), so S-cases in earlier weeks include other non-VOC lineages, especially outside the East and South East of England. Plots for all STP areas are shown in Figure S3.
Transmission advantage of the VOC
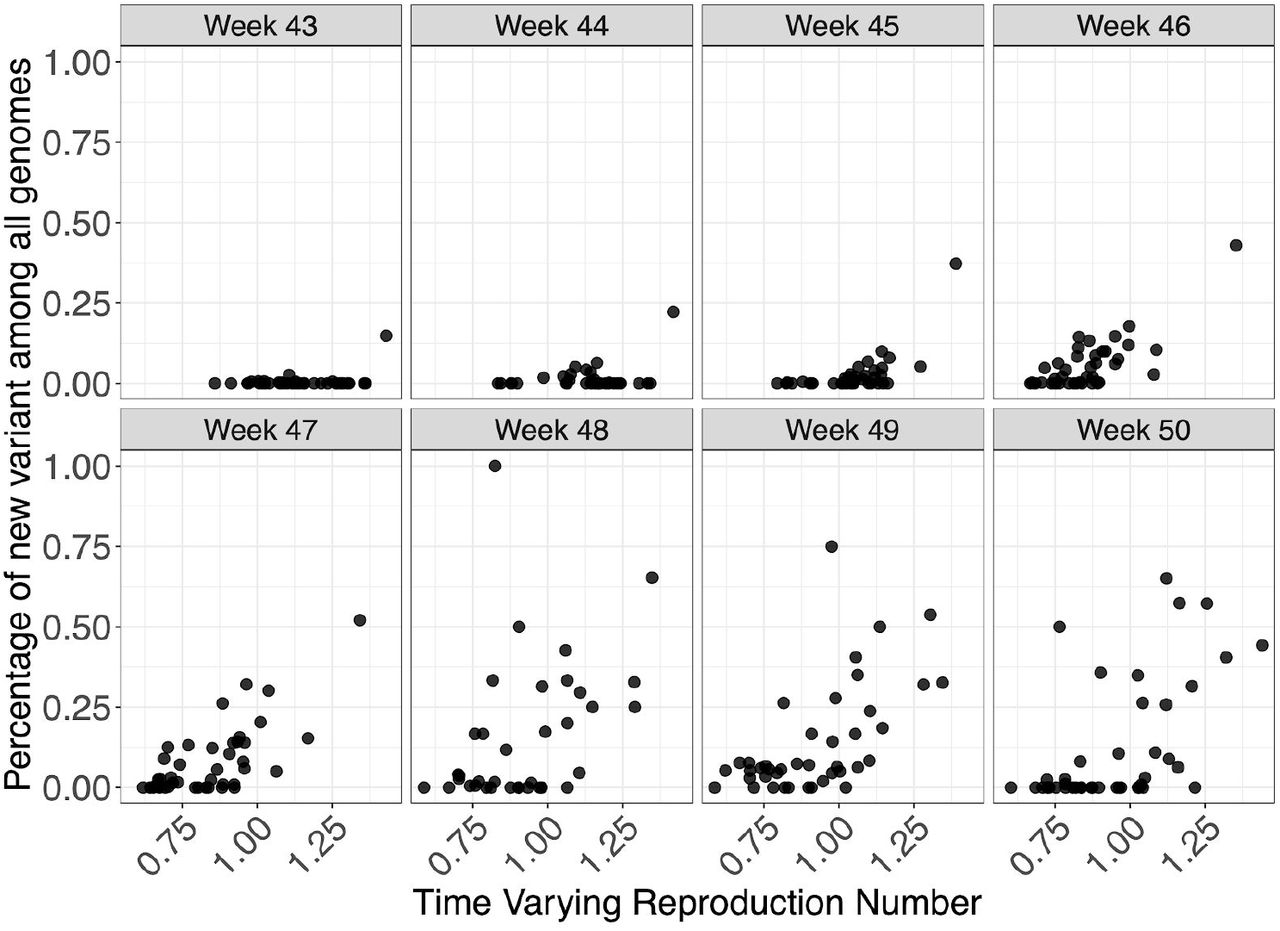
To examine the differences between S- and S+ growth rates, we focus on epidemiological weeks 46-50 (8th November-12th December). We estimate the total S- and S+ in each STP and week by adjusting counts upwards in proportion to total cases reported in each STP and week. We then calculate the week on week growth factor in both S- and S+ cases by dividing the case numbers in week t+1 by the case numbers in week t. Given an assumed mean generation time of SARS-CoV-2 of 6.5 days9, we correct these weekly growth factors by raising them to the power of 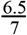 to ensure they can be interpreted as approximate reproduction numbers. For each STP and week, we compute both the ratio and difference of the resulting empirical reproduction number of the S-negative cases to that of the S-positive cases (Figure 3). Overall, the median multiplicative advantage is 1.74 for the VOC, and the median additive advantage is 0.63, showing a clear advantage of the VOC for both metrics.
to ensure they can be interpreted as approximate reproduction numbers. For each STP and week, we compute both the ratio and difference of the resulting empirical reproduction number of the S-negative cases to that of the S-positive cases (Figure 3). Overall, the median multiplicative advantage is 1.74 for the VOC, and the median additive advantage is 0.63, showing a clear advantage of the VOC for both metrics.

Empirical data analysis of the advantage in weekly growth factors (cases in week t+1 divided by cases in week t) for the VOC versus non-VOC lineages. Each point represents either the ratio (left) or difference (right) of weekly growth factors for the VOC versus non-variant for an NHS England STP area and week, using the raw SGTF data shown in Figure S1 (not correcting for TPR). Colours and shapes differentiate epi weeks. Numbers above 1 on the top plot and above 0 on the bottom plot show a transmission advantage. The blue line represents the mean advantage for a particular proportion of VOC among all cases, and the grey lines the 95% envelope. Scatter at low frequencies largely reflects statistical noise due to low counts.
Paired growth rate trends of the VOC and non-VOC lineages demonstrate an increase in the reproduction number
We next tested the hypothesis that the higher growth rates of the VOC compared to other circulating lineages might be due solely to shorter generation times (e.g. a shorter incubation period), rather than increased transmissibility (R). To this end, we compared the number of NHS STP areas in which both VOC and non-VOC cases increased or decreased (Table 1). If the VOC had the same reproduction number as non-VOC but a shorter generation time, VOC cases are expected to grow faster than non-VOC cases in areas where non-VOC grew. However VOC cases are expected to decline faster than non-VOC cases where non-VOC declined. Furthermore, areas where VOC grew but non-VOC declined would, on average, be equally balanced by areas where the opposite was true. That is, if only the generation interval of the VOC had shortened, the proportion of areas with positive growth of the VOC and negative growth of the non-VOC would be highly correlated with the proportion of areas with negative growth of the VOC and positive growth of the non-VOC. However, of 168 STP-weeks (42 STP areas, weekly growth factors for weeks 46-49) there were 97 STP-weeks where growth was observed in S- and decline was observed in S+, but only 1 STP-week where the opposite was true (Table 1), indicating strong evidence against S+ and S-reproduction numbers being equal (McNemar’s Chi-square test with continuity correction test statistic 92.02, p < 1e-15). Comparing the empirical distribution of growth factors from S+ and S-with the nonparametric Kolmogorov–Smirnov test results in rejecting the null hypothesis (p < 1e-15) that the two arise from the same probability distribution.
Contingency table of VOC and non-VOC weekly growth factors derived from raw SGTF data within 42 NHS STP areas for weeks 46-49, stratified by increasing (>1) and declining incidence(≤ 1). The imbalance in off-diagonal elements gives strong evidence of increased transmissibility, even if the VOC had an altered generation time distribution.
Share of age groups among VOC and non-VOC cases
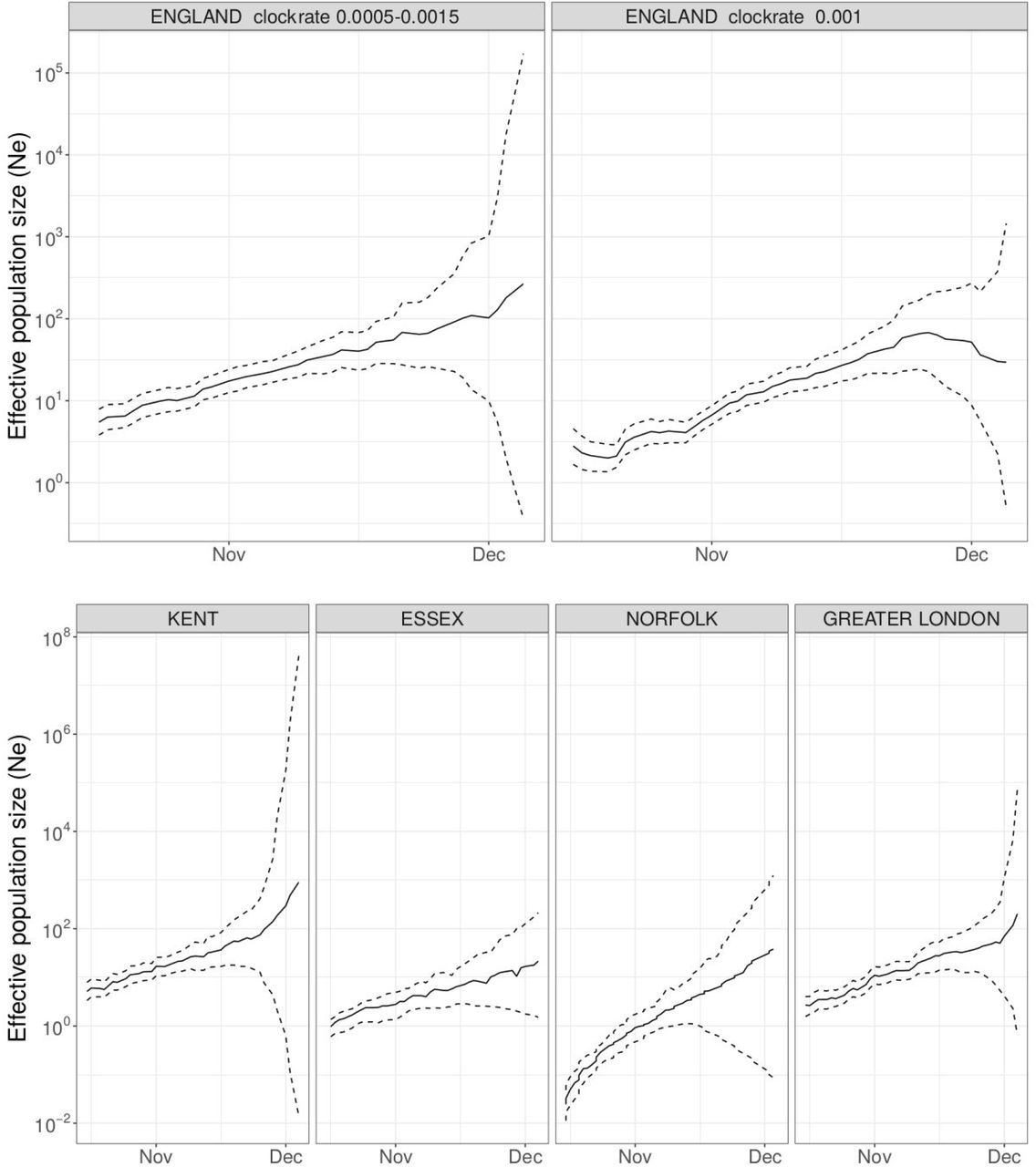
To assess differences in the age distribution of VOC versus non-VOC cases, we considered S- and S+ case numbers in weeks 46-51 across NHS STP regions. Case numbers were standardised for differences in the population age composition in each area, weighted to compare S-cases from each NHS STP region and each epidemiological week with an equal number of S+ cases from that same STP and week (a case-control design), and aggregated over STP weeks. Accounting for binomial sampling variation and variation by area and week, we observe significantly more S-cases, our biomarker of VOC cases, among individuals aged 0-19 as compared to S+ cases, and significantly fewer S-cases among individuals aged 60-79 (Figure 4). This trend is seen in each of the regions of England most affected by the VOC thus far (East of England, London, South East and Midlands), and similar differences are seen between the raw (non-case control weighted, and non-age-standardised) age distributions of S+ and S-cases.

Age distribution of S-gene negative (S-) and S-gene positive (S+) PCR-positive pillar 2 cases from the SGSS dataset (not adjusted for TPR). Case numbers are weighted to compare S-cases from each NHS STP region and epidemiological week with an equal number of S+ cases from that STP and week (a case-control design), and standardised for differences in the age composition of each STP area. (A) Age distribution of S- and S+ cases. (B) Ratio of S- to S+ proportions of cases in each 10 year band. Results shown are for weeks 46-51. Ages were capped at 80. 95% empirical confidence intervals calculated by bootstrapping over STP areas and weeks, and sampling variation within STP areas and weeks.
Regression analysis of VOC transmissibility
To investigate the effect of VOC frequency on the overall time-varying reproduction number, Rt, we undertook a number of regression analyses. We conduct our analyses at two different spatial scales - lower tier local authority (LTLA) and NHS STP areas. For each, we estimated Rt by week and area using data on pillar 2 testing, deaths and hospitalisations using a previously described model9,11. Figure 5 shows the empirical relationship between weekly estimates of Rt at STP level and the frequency of the VOC estimates using genomic data.

Relationship between genomic frequency of the VOC lineage among all genomes plotted against the time varying reproduction number for each week. Each datapoint is an STP area.
We apply a range of frequentist models with a bootstrapping procedure to account for non-normality in responses, as well as a Bayesian regression which explicitly models VOC frequency, such that it simultaneously informs the parameter for binomially-distributed observations of frequency and the Rt estimates. The role of geography in explaining variance of Rt was examined using both fixed and random effects. These models were applied to both genomic-based frequency estimates and TPR-adjusted SGTF proportions of pillar 2 cases for which S-gene data was available. Given this definition and the approximately 1 week generation time of SARS-CoV-2, we expect Rt to have stronger association with VOC frequency 1 week earlier. We therefore present regressions of Rt against frequency at week t-1 for our default analysis (where t spans weeks 44-50), and a regression of Rt against frequency at week t is provided in the Supplementary Information.
Regression results are reported in Table 2 (Table S2 for sensitivity analysis). We estimate the additive effect on Rt, i.e., the increase or decrease in Rt (using Rt as response in the linear model) due to the variant. As an example, with an additive effect size of 0.4, an area with an Rt of 0.8 without the VOC would have an Rt of 1.2 if only the VOC was present. As expected, models which allow for fixed effects of week and region give lower effect sizes for the VOC than random effect models, given the latter constrain week and time effects more than fixed effect models, due to the assumptions that such effects arise from normal distributions. The Bayesian model results closely resemble those from the frequentist random effects model.
Estimated additive change of reproduction numbers of VOC compared with other variants for different regression models, spatial resolutions, and data used to estimate the prevalence of the VOC. Analysis uses Rt estimates from weeks 44-50 and data on the proportion of the VOC one week earlier, to take account of the generation time of SARS-CoV-2.
The results in Table 2 show a clear association between the VOC and Rt. However, this analysis cannot prove causality. The estimated additive effect is specific to the conditions that prevailed in England during the time period examined.
Estimating reproduction numbers for VOC and non-VOC independently
We estimated the reproduction number of the VOC via phylodynamic analysis of whole genome sequences from Pillar 2 national SARS-CoV-2 testing, sampled up to December 6, 2020. First, we fitted a non-parametric skygrowth model 12 by maximum likelihood to 776 genomes that we selected from England in inverse proportion to the number of diagnosed cases sequenced in each region by week (see Supporting Methods). This model indicates that the effective population size of VOC 202012/01 grew at a relatively stable rate of 58% per week from September 20 to December 6, corresponding to a reproduction number of 1.59. Estimates of growth rate were insensitive to uncertainty in the molecular clock rate of evolution. Second, we fitted the model to genomes from four regions with more than fifty sequences, Kent (n=701), Greater London (n=606), Essex (n=131), and Norfolk (n=81). This regional analysis indicated growth rates ranging from 58% to 92% per week, corresponding to reproduction numbers between 1.56 and 1.95 (Figure S6). Finally, we carried out a Bayesian non-parametric coalescent analysis using the Skygrid model13 using the same set of 776 genomes. This analysis showed growth until the start of November followed by a plateau for the month of November coincident with the second English lockdown (Figure S7). This suggests the lockdown constrained growth of the VOC, but was insufficient to cause a reduction in incidence. To estimate parameter values we also estimated the initial growth rate of the VOC lineage under a parametric logistic growth coalescent model14. Under this model we estimated a growth rate of 71.5 per year, corresponding to a doubling time of 3.7 days (95% CrI: 2.4 – 4.9) and a reproduction number of 2.27 (1.84 – 2.73). By comparison, a simple exponential growth model over this entire period yields a growth rate of 27.9 with a doubling time of 9.1 days (7.4, 11.2) and reproductive number of 1.50 (1.40 – 1.60).
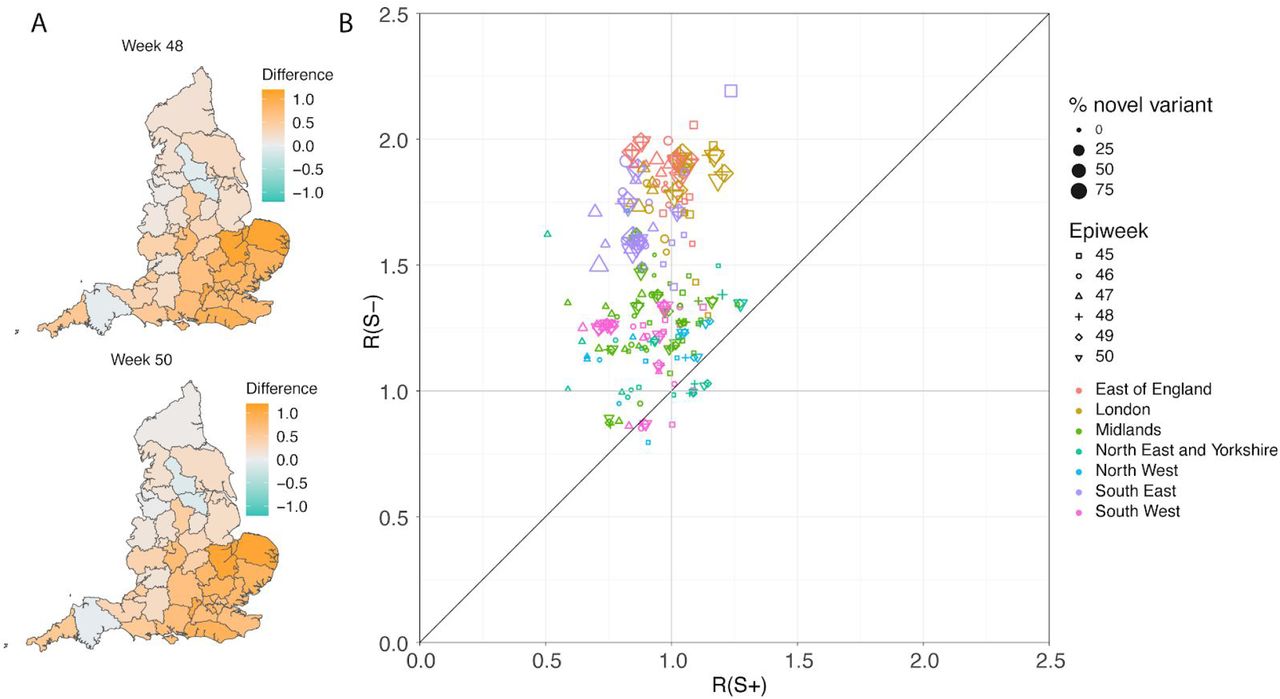
In a parallel epidemiological analysis, we estimated VOC and non-VOC pillar 2 case numbers by STP area using TPR-corrected SGTF frequencies applied to overall PHE pillar 2 case numbers. We then estimate Rt by week separately for VOC and non-VOC, using the same model previously used to generate overall (non lineage-stratified) Rt estimates11. We first fit the unstratified model to estimate the infection ascertainment ratio (numbers of infections being identified as positive cases) and infection seeding (initial infections in each region). For seeding, we use the estimated infections from our unstratified model. The mean number of daily infections for week 42 and 43 are used for seeding both VOC and non-VOC models. The fraction of SGTF cases is used to distribute infections for seeding between VOC and non-VOC in weeks 42 and 43. We then compute Rt estimates for weeks 45-50, to avoid the seeding assumptions affecting Rt estimates. Figure 6A shows the mean posterior difference between Rt estimates for VOC and non-VOC for week 48 and 50, while figure 6B shows plots median Rt estimates for VOC and non-VOC across all NHS regions for weeks 45-50. The Rt estimates for VOC are greater than those for non-VOC for 94% of STP-week pairs (points above the diagonal in Figure 6B). Figure S4 shows the mean posterior difference between Rt estimates for VOC and non-VOC for all weeks 45-50, while Figure S5 shows the ratio of Rt estimates. The mean Rt difference across weeks 45-50 is 0.51 [95% CrI: −0.09 − 1.10] which was computed from the set of 42×6 (STP x week) posteriors of Rt estimated for the VOC and non-VOC. The mean ratio of the estimated Rt for the VOC and non-VOC was 1.56 [95%CI: 0.92 − 2.28] for the same period, see Figure S5. Aggregating across all STPs we find that the mean Rt during the second English lockdown across all STPs was 1.45 [0.91-1.89] for the VOC and 0.92 [0.86-1.06] for non-VOC strains.

(A) Map of the difference in median Rt estimates for VOC and non-VOC variants for all STPs for weeks 48 and week 50. (B) Scatterplot of the reproduction numbers of VOC (S-) and non-VOC (S+) by STP and week. Point size indicates frequency of the VOC, while shape and colour signify week and NHS region, respectively.
Discussion
While evidence has accumulated that substitutions associated with the B.1.1.7 lineage are associated with significant changes in virus phenotype2–4,15, assessing the extent to which these changes lead to meaningful differences in transmission between humans is challenging and cannot be evaluated experimentally. When randomised experimental studies are not possible, observational studies provide stronger evidence if consistent patterns are seen in multiple locations and at multiple times. While rapidly increasing frequency of a new lineage within a viral population is consistent with a selective advantage, it is also possible that increases in frequency may be caused by founder effects or genetic drift, especially for genetic variants which are repeatedly introduced from overseas16,17. But in contrast to previous genetic variants which have achieved high prevalence, we see expansion of the VOC from within the United Kingdom and a pattern of faster epidemic growth in tandem with expansion of the VOC has been repeated in multiple regions. In this paper we have focussed on spatiotemporally stratified analyses using a variety of statistical approaches to evaluate the relationship between SARS-CoV-2 transmission intensity and the frequency of the VOC, B.1.1.7 during November-December 2020 in different UK regions.
Assessment of the transmission characteristics of the VOC (B.1.1.7) was aided by the high correlation between its frequency and the occurrence of S-gene target failure (SGTF) in routine PCR testing of community cases of COVID-19 associated with the Δ69-70 deletion present in the VOC lineage (Figure 1 and S1). S-gene positivity results were available for over a third of all PCR-positive community COVID-19 cases for November and December 2020, allowing us to use SGTF frequency as a proxy for VOC frequency, and thus estimate VOC and non-VOC incidence trends by region over that time period. We see a very clear visual association between SGTF frequency and epidemic growth in nearly all areas (Figures 2 and S3), which is reinforced by empirical assessment of area-specific week on week growth factors of VOC and non-VOC case numbers (Figure 3) and by formal regression analyses of the association between estimates of local Rt and VOC frequency estimated from SGTF data (Table 2).
Finally, we used the SGTF data to independently estimate Rt by region and week for the VOC and non-VOC variants (Figures 6 and S4) and derived similar estimates for the increase in Rt associated with the VOC. This latter analysis is perhaps the most powerful, as no parametric assumptions are made about the relationship between Rt of the VOC and that of non-VOC strains.
Phylodynamic modelling provides additional information about growth of the VOC in October during a period when SGTF data is sparse. Although not apparent in all analyses, this suggests that the VOC expanded rapidly in October, with growth slowing (but not reversing) during national lockdown in November (Figures S6 and S7).
We were also able to rule out the hypothesis that increased incidence growth rates in the VOC are solely due to a change in the latent period or generation time distribution, but not the reproduction number itself (Table 1), since we see a large and statistically significant imbalance between regions where the VOC increased and where the non-VOC decreased, and vice-versa. A change solely in, for instance, the latent period would not be expected to change the direction of incidence growth.
We quantified the transmission advantage of the VOC relative to non-VOC lineages in two ways: as an additive increase in R that ranged between 0.4 and 0.7, and alternatively as a multiplicative increase in R that ranged between a 50% and 75% advantage. We were not able to distinguish between these two approaches in goodness-of-fit, and either is plausible mechanistically. A multiplicative transmission advantage would be expected if transmissibility had increased in all settings and individuals, while an additive advantage might reflect increases in transmissibility in specific subpopulations or contexts. More generally, the temporal context is important; these estimates of transmission advantage apply to a period where high levels of social distancing were in place in England; extrapolation to other transmission contexts, without detailed knowledge of the drivers of transmission, requires caution.
We observe a small but statistically significant shift towards under 20s being more affected by the VOC than non-VOC variants (Figure 4), even after controlling for variation by week and region. However, as with our earlier results, this observation does not resolve the mechanism that might underlie these differences. Differences between the age-distributions of VOC and non-VOC community cases may result from the overall increase in transmissibility of the VOC (especially during a time where lockdown was in force but schools were open), increased susceptibility of under 20s, or more apparent symptoms (and thus a propensity to seek testing) for the VOC in that age range.
There are a number of limitations to our analysis. The genomic and epidemiological data analysed was collected as part of routine surveillance, and thus may not be an entirely representative sample of SARS-CoV-2 infections in England over the time period considered. We also focussed on relatively simple, data-driven analyses using relatively simple models making parsimonious assumptions, rather than, for instance, attempting to model the long-term transmission dynamics of VOC and non-VOC lineages more mechanistically. We also did not attempt to explicitly model the spatiotemporal correlation intrinsic in infectious disease data, especially when considering the spread of a new variant from a point source. Doing so is an important priority for future work, but will require explicit incorporation of data on population movement patterns.
Early versions of our analyses informed the UK government policy response to this VOC and that of other countries. The substantial transmission advantage we have estimated the VOC to have over prior viral lineages poses major challenges for ongoing control of COVID-19 in the UK and elsewhere in the coming months. Social distancing measures will need to be more stringent than they would have otherwise. A particular concern is whether it will be possible to maintain control over transmission while allowing schools to reopen in January 2021. These policy questions will be informed by the ongoing urgent epidemiological investigation into this variant, most notably examining evidence for any changes in severity, but also giving more nuanced understanding into transmissibility changes, for instance in the household setting.
Data Availability
All aggregated data to reproduce analysis will be provided in the url below.
Funding
COG-UK is supported by funding from the Medical Research Council (MRC) part of UK Research & Innovation (UKRI), the National Institute of Health Research (NIHR) and Genome Research Limited, operating as the Wellcome Sanger Institute. The Imperial College COVID-19 Research Fund, UKRI (MR/V038109/1), The Academy of Medical Sciences (SBF004/1080), Bill & Melinda Gates Foundation (OPP1197730, OPP1175094), the European Commission (CoroNAb 101003653), the NIHR BRC Imperial College NHS Trust Infection and COVID themes (RDA02), Amazon AWS and Microsoft AI for Health, the EPSRC, The Medical Research Council (MR/R015600/1), the NIHR Health Protection Research Unit for Modelling and Health Economics, NIHR VEEPED project funding (PR-OD-1017-20002). Wellcome core funding to the Wellcome Sanger Institute (206194). JTM, NL and AR acknowledge the support of the Wellcome Trust (Collaborators Award 206298/Z/17/Z – ARTIC network). AR is supported by the European Research Council (grant agreement no. 725422 – ReservoirDOCS).
Supplementary Information
Methods
All code and data is available at https://github.com/mrc-ide/covid19-variant-N501Y
Logistic growth model applied to variant frequency data
The logistic growth model fitted to VOC frequency data arises from a simple mechanistic model for competition between two strains. Let established lineages have a reproduction number R and let the VOC have reproduction number R(1+s). According to this model, the log odds of observing a variant over time will be proportional to (R/g)st, where R is assumed to be constant over weeks 44-49, g is the generation time, s is a selection coefficient (assumed to be constant) and t is time. If these conditions are met, s can be interpreted as a multiplicative change in the reproduction number or as a change in the generation time or as some combination of these factors. From available data on times of sampling each variant, the compound parameter (R/g)s can be estimated. An approximate estimate of s is obtained by treating R =1 and generation time g=6.5 days as a constant for weeks 44-49. In the text, we refer to s as the change in growth per generation, and is comparable to multiplicative changes in Rt estimated using other methods.
Phylodynamic analysis
Analysis was based on Pillar 2 whole genome sequences with known Upper Tier Local Authority (UTLA). Maximum likelihood non-parametric phylodynamic analysis was carried out by: 1) estimating a maximum likelihood phylogeny in IQtree18 (HKY model of sequence evolution); 2) We removed duplicated identical sequences and estimated a time-scaled phylogeny using treedater19 using a strict molecular clock. The molecular clock rate of evolution was constrained to 0.0005 − 0.0015 substitutions per site per year. Small branch lengths in the tree were collapsed and polytomies randomly resolved to produce 20 new variations on the dated ML tree. 3) The skygrowth model was then fitted to these dated trees with parameters 64 time steps and tau bounded between 0.0025-20. Growth rate estimates were translated to reproduction numbers using the method of Wallinga and Lipsitch20 in the epitrix R package and using serial intervals from Flaxman et al.9 The main text reports the median growth rate over time. A sensitivity analysis was carried out with the molecular clock rate fixed at 0.001 substitutions per site per year which yielded only marginally different estimates of the median growth rate.
Sequence sample weights were used to select samples for phylodynamic modelling. Weights, assigned to sequence samples according to their UTLA and their collection date, correspond to the number of confirmed cases represented by each sequence in a UTLA relative to other UTLAs on the same date. To smooth over sparsity, case and sample counts were summed over the fourteen days prior to the date. Confirmed cases were accessed from the ONS API (https://api.coronavirus.data.gov.uk). Code to compute sequence sample weights is available at https://github.com/robj411/sequencing_coverage.
Bayesian coalescent phylodynamic analysis was performed using BEAST v1.10.414 employing the Skygrid non-parametric approach 21 and exponential and logistic growth parametric models. A Jukes-Cantor model of substitution and a strict molecular clock were assumed. MCMC chains were run for 100M states, removing 10M as ‘burn-in’ and then thinning to 9000 samples from the posterior. Parameters were summarised and demographic curves reconstructed using Tracer v1.7.2 22.
SGTF as a biomarker for the VOC and frequency of SGTF over time
Data on SGTF among pillar 2 tests was obtained from the 3 largest (“Lighthouse”) PCR testing laboratories and integrated into the PHE Second Generation Surveillance System (SGSS) database. We also obtained the total number of cases reported by Public Health England.
Application of SGTF as a diagnostic for the VOC provides a large advantage over genomic sequencing in terms of cost, speed, and the sample size of available test results. We extracted 275,571 S target positive (S+) and 96,070 S target negative (S-) test results collected between 1 October and 19 December, 2020 and examined the potential to use SGTF cases (S-) as a biomarker for the VOC lineage. While the tests are not a representative sample of infections over this time period, they are a representative sample of tests within a given region and week and thus provide information about the relative abundance of the VOC versus other variants over time and between regions.
Other lineages have been observed to carry Δ69-70 which is associated with SGTF and which would have a similar impact on the TaqPath assay. The diagnostic specificity of SGTF will therefore vary over time and space as it depends on the abundance of other lineages with Δ69-70. This deletion is mostly found in global lineage B.1.258 and is highly linked with the spike N439K variant23. Lineage B.1.258 has circulated in the UK since June 2020 where it is now widespread. However, the frequency of this variant has been relatively stable since October 2020.
In order to capture diagnostic uncertainty with SGTF, we fitted a spatio-temporal model of the frequency of the VOC relative to other variants carrying Δ69-70. We fitted a generalized additive model24,25 to counts of genomes classified 26 as belonging to the VOC lineage or genomes which do not belong to the VOC lineage but still carry Δ69-70. Counts were tabulated by LTLA and epi week. We used a cubic spline to model trends over time, and correlation between neighbouring LTLAs was modeled with a Gaussian Markov Random Field (GMRF). This model was used to predict the true positive rate (TPR) − the probability that a sample collected at a particular time and place belongs to the VOC lineage, given that it carries Δ69-70. Figure S1 shows how this prediction has changed over weeks spanning November 2020. From week 50 onwards, we assumed the TPR was 1 across England. The fitted model thus provided an estimate of the true positive rate (TPR) of using SGTF as a proxy for VOC frequency as a function of time and region in England (Figure S2).
Regression analysis of VOC transmissibility
Using reported COVID-19 PCR-positive case counts and deaths, we estimated the time-varying reproduction numbers Rt for epidemiological weeks 44-50 in each LTLA and STP in England. These estimates were obtained from a previously developed11 Bayesian semi-mechanistic transmission model with a latent weekly random walk process that does not include underlying factors that drive transmission. The STP model is based on pillar 2 cases only, and includes hospital admissions as an additional input; the LTLA model is based on both pillar 1 and pillar 2 data. Further model information can be found in previous publications9,27. These Rt estimates refer to the reproduction number of the infections that gave rise to the infections at time t. Given this definition and the approximately 1 week generation time of SARS-CoV-2, we would expect Rt to be most closely associated with VOC frequency 1 week earlier. We therefore present regressions of Rt against frequency at week t-1 for our default analysis (where t spans weeks 44-50), but present results for regression of Rt against frequency at week t as a sensitivity analysis in Supplementary Information.
We consider two data sources when estimating the proportion of the VOC: Genomic-based frequency estimates and TPR-adjusted SGTF proportions of pillar 2 cases for which S-gene data was available. For the SGTF-based estimates, frequency estimates were available for all STPs and LTLAs for the weeks 43-49 (294 STP-weeks in total). When using genomic data, 7 STP-weeks in the week range 43-49 had no sampled genomes, leaving 287 STP-weeks for analysis, for which we had an average of 137 genomes per STP week (of which on average 7 were the VOC).
We use two types of frequentist models. The first uses a fixed effect for each area, the second uses a random effect for each area. Fixed effects of epidemiological week were included in both cases. Confidence intervals for the fixed and random effects models were computed through a bootstrapping method which resamples the areas with replacement and also samples the counts of VOC in each week/area pair based on the observed counts. This bootstrap aims to account for potential skewness and non-normality of responses, dependence within areas and the randomness in the proportion of the VOC sampled within an area. When using genomic-based frequency estimates we excluded STP-weeks that had fewer than 5 genetic profiles, as these were deemed to not give reliable estimates of the proportion of VOC. This removed a further 14 out of the 287 STP-weeks from those analyses.
We also implemented a Bayesian regression model 28 at STP level, taking into account uncertainty in the frequency estimates and uncertainty in the Rt estimates. VOC frequency was modelled explicitly, such that it simultaneously informed the parameter for binomially-distributed observations of frequency and the Rt estimates. The regression for Rt included terms for the VOC frequency, the week (modelled with a smooth spline) and the area as a factor. The formula to describe the binomial frequency data includes a linear function of time with an area-dependent slope, and was fitted to the genome counts and the counts of S-negative cases separately.
Estimated growth difference per generation of B.1.1.7 by English administrative region based on a maximum likelihood estimation of logistic growth in variant frequency and assuming a 6.5 day generation time and Rt=1 for established lineages. Confidence intervals are based on likelihood profiles
Estimated additive change of reproduction numbers of VOC compared with non-VOC using different regression models, spatial resolutions, and data to estimate the prevalence of the VOC. Analyses use data on the proportion of the VOC and estimates of Rt in weeks 44-50.

Estimate of true positive rates for classification of B.1.1.7 infection given SGTF result (S-) as a function of time and UK region. The colour gradient shows the probability of sampling a B.1.1.7 sequence conditional on sampling any sequence with Δ69-70.

Empirical (point) and estimated (line) frequencies of TPR-adjusted SGTF frequencies over time. The size of points correspond to the number of samples observed by day. Confidence intervals show 95% estimated sampling error for daily proportions.

Case trends in all NHS STP areas, ordered by decreasing frequency of S-in the last week shown. Total cases reported are shown as a thick line. A subset of these - those tested in the 3 largest “Lighthouse” laboratories - were tested for SGTF. The total cases line is coloured according to percentage S-among those tested. Counts of S+ and S-reported via the PHE SGSS system are shown by the thin lines. The dates of the second lockdown are indicated by the vertical red lines. Raw SGTF data are shown here (not adjusted for TPR), so S-cases in earlier weeks include other non-VOC lineages, especially outside the East and South East of England.

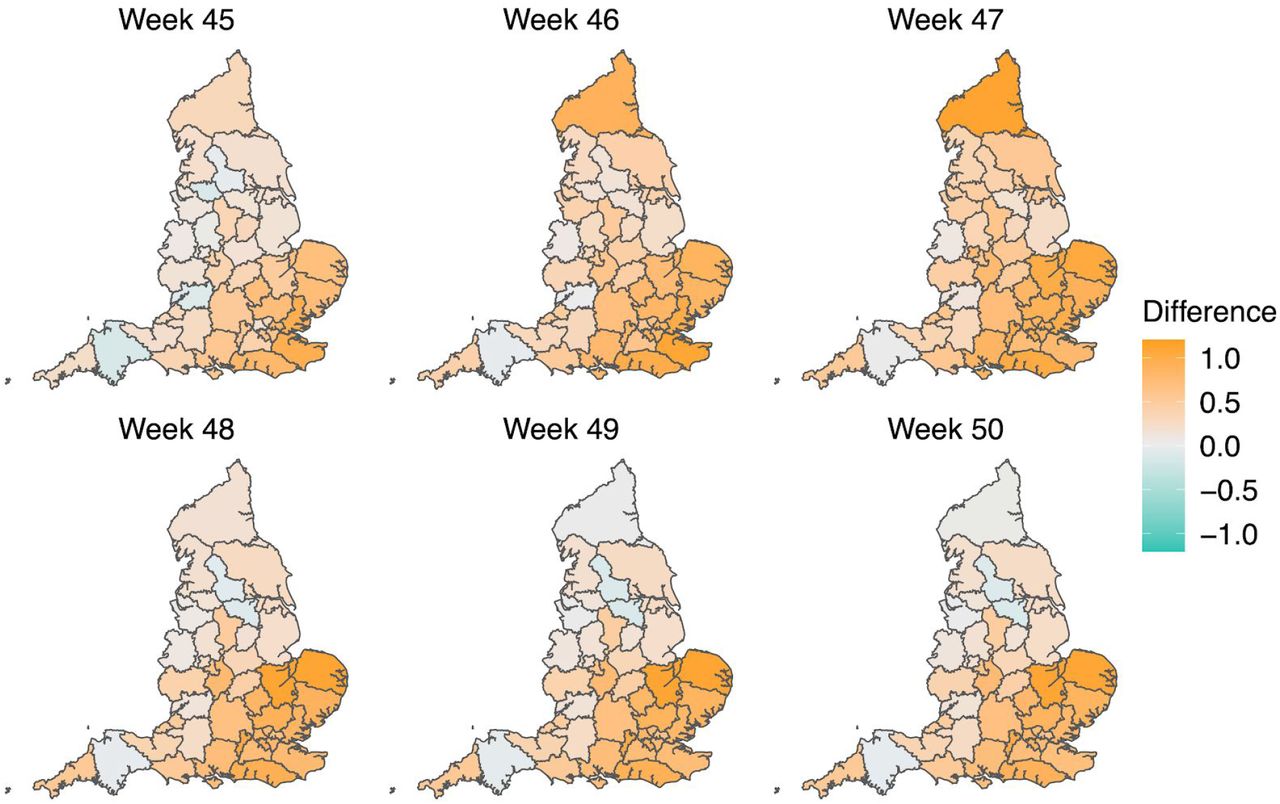
Map of the difference in median Rt estimates for VOC and non-VOC variants for all STPs between week 45 to week 50. The darker orange color indicates the additive advantage VOC has over non-VOC variant for Rt, whereas the darker green color shows the advantage for non-VOC variant over VOC.

Map of the ratio of median Rt estimates for VOC and non-VOC variants for all STPs between week 45 to week 50. Darker color indicates the higher multiplicative advantage for VOC variant in comparison to the non-VOC variant. The mean of the ratio between R estimates for S- and S+ for all posterior samples across weeks 45 to 50 and all STPs is 1.56, with 95% CI [0.92 − 2.28]1.
Maximum likelihood skygrowth estimates of the effective population size of the VOC through time in England and in four areas of England with more than 80 whole genome sequences. In the upper right panel the molecular clock rate of evolution was fixed at 0.001 substitutions per site per year. In all other analyses the rate was estimated and bound between 0.0005 and 0.0015 substitutions per site per year.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Bayesian estimates of effective population size through time based on 776 genomes sampled between October and December 6. Top and bottom panels show estimates based on a Bayesian skygrid model overlaid with fits of different parametric models. A) Estimates based on a logistic growth model. B) Estimates from an exponential growth model.
Supplementary Appendix: COG-UK consortium members
Funding acquisition, Leadership and supervision, Metadata curation, Project administration, Samples and logistics, Sequencing and analysis, Software and analysis tools, and Visualisation: Dr Samuel C Robson 13.
Funding acquisition, Leadership and supervision, Metadata curation, Project administration, Samples and logistics, Sequencing and analysis, and Software and analysis tools:
Prof Nicholas J Loman 41, Dr Thomas R Connor 10, 69.
Leadership and supervision, Metadata curation, Project administration, Samples and logistics, Sequencing and analysis, Software and analysis tools, and Visualisation:
Dr Tanya Golubchik 5.
Funding acquisition, Metadata curation, Samples and logistics, Sequencing and analysis, Software and analysis tools, and Visualisation:
Dr Rocio T Martinez Nunez 42.
Funding acquisition, Leadership and supervision, Metadata curation, Project administration, and Samples and logistics:
Dr Catherine Ludden 88.
Funding acquisition, Leadership and supervision, Metadata curation, Samples and logistics, and Sequencing and analysis:
Dr Sally Corden 69.
Funding acquisition, Leadership and supervision, Project administration, Samples and logistics, and Sequencing and analysis:
Ian Johnston 99 and Dr David Bonsall 5.
Funding acquisition, Leadership and supervision, Sequencing and analysis, Software and analysis tools, and Visualisation:
Prof Colin P Smith 87 and Dr Ali R Awan 28.
Funding acquisition, Samples and logistics, Sequencing and analysis, Software and analysis tools, and Visualisation:
Dr Giselda Bucca 87.
Leadership and supervision, Metadata curation, Project administration, Samples and logistics, and Sequencing and analysis:
Dr M. Estee Torok 22, 101.
Leadership and supervision, Metadata curation, Project administration, Samples and logistics, and Visualisation:
Dr Kordo Saeed 81, 110 and Dr Jacqui A Prieto 83, 109.
Leadership and supervision, Metadata curation, Project administration, Sequencing and analysis, and Software and analysis tools:
Dr David K Jackson 99.
Metadata curation, Project administration, Samples and logistics, Sequencing and analysis, and Software and analysis tools:
Dr William L Hamilton 22.
Metadata curation, Project administration, Samples and logistics, Sequencing and analysis, and Visualisation:
Dr Luke B Snell 11.
Funding acquisition, Leadership and supervision, Metadata curation, and Samples and logistics:
Dr Catherine Moore 69.
Funding acquisition, Leadership and supervision, Project administration, and Samples and logistics:
Dr Ewan M Harrison 99, 88.
Leadership and supervision, Metadata curation, Project administration, and Samples and logistics:
Dr Sonia Goncalves 99.
Leadership and supervision, Metadata curation, Samples and logistics, and Sequencing and analysis:
Prof Ian G Goodfellow 24, Dr Derek J Fairley 3, 72, Prof Matthew W Loose 18 and Joanne Watkins 69.
Leadership and supervision, Metadata curation, Samples and logistics, and Software and analysis tools:
Rich Livett 99.
Leadership and supervision, Metadata curation, Samples and logistics, and Visualisation:
Dr Samuel Moses 25, 106.
Leadership and supervision, Metadata curation, Sequencing and analysis, and Software and analysis tools:
Dr Roberto Amato 99, Dr Sam Nicholls 41 and Dr Matthew Bull 69.
Leadership and supervision, Project administration, Samples and logistics, and Sequencing and analysis:
Prof Darren L Smith 37, 58, 105.
Leadership and supervision, Sequencing and analysis, Software and analysis tools, and Visualisation:
Dr Jeff Barrett 99 and Prof David M Aanensen 14, 114.
Metadata curation, Project administration, Samples and logistics, and Sequencing and analysis:
Dr Martin D Curran 65, Dr Surendra Parmar 65, Dr Dinesh Aggarwal 95, 99, 64 and Dr James G Shepherd 48.
Metadata curation, Project administration, Sequencing and analysis, and Software and analysis tools:
Dr Matthew D Parker 93.
Metadata curation, Samples and logistics, Sequencing and analysis, and Visualisation:
Dr Sharon Glaysher 61.
Metadata curation, Sequencing and analysis, Software and analysis tools, and Visualisation: Dr Matthew Bashton 37, 58, Dr Anthony P Underwood 14, 114, Dr Nicole Pacchiarini 69 and Dr Katie F Loveson 77.
Project administration, Sequencing and analysis, Software and analysis tools, and Visualisation:
Dr Alessandro M Carabelli 88.
Funding acquisition, Leadership and supervision, and Metadata curation:
Dr Kate E Templeton 53, 90.
Funding acquisition, Leadership and supervision, and Project administration:
Dr Cordelia F Langford 99, John Sillitoe 99, Dr Thushan I de Silva 93 and Dr Dennis Wang 93.
Funding acquisition, Leadership and supervision, and Sequencing and analysis:
Prof Dominic Kwiatkowski 99, 107, Prof Andrew Rambaut 90, Dr Justin O’Grady 70, 89 and Dr Simon Cottrell 69.
Leadership and supervision, Metadata curation, and Sequencing and analysis:
Prof Matthew T.G. Holden 68 and Prof Emma C Thomson 48.
Leadership and supervision, Project administration, and Samples and logistics:
Dr Husam Osman 64, 36, Dr Monique Andersson 59, Prof Anoop J Chauhan 61 and Dr Mohammed O Hassan-Ibrahim 6.
Leadership and supervision, Project administration, and Sequencing and analysis:
Dr Mara Lawniczak 99.
Leadership and supervision, Samples and logistics, and Sequencing and analysis:
Prof Ravi Kumar Gupta 88, 113, Dr Alex Alderton 99, Dr Meera Chand 66, Dr Chrystala Constantinidou 94, Dr Meera Unnikrishnan 94, Prof Alistair C Darby 92, Prof Julian A Hiscox 92 and Prof Steve Paterson 92.
Leadership and supervision, Sequencing and analysis, and Software and analysis tools:
Dr Inigo Martincorena 99, Prof David L Robertson 48, Dr Erik M Volz 39, Dr Andrew J Page 70 and Prof Oliver G Pybus 23.
Leadership and supervision, Sequencing and analysis, and Visualisation:
Dr Andrew R Bassett 99.
Metadata curation, Project administration, and Samples and logistics:
Dr Cristina V Ariani 99, Dr Michael H Spencer Chapman 99, 88, Dr Kathy K Li 48, Dr Rajiv N Shah 48, Dr Natasha G Jesudason 48 and Dr Yusri Taha 50.
Metadata curation, Project administration, and Sequencing and analysis:
Martin P McHugh 53 and Dr Rebecca Dewar 53.
Metadata curation, Samples and logistics, and Sequencing and analysis:
Dr Aminu S Jahun 24, Dr Claire McMurray 41, Ms Sarojini Pandey 84, Dr James P McKenna 3, Dr Andrew Nelson 58, 105, Dr Gregory R Young 37, 58, Dr Clare M McCann 58, 105 and Mr Scott Elliott 61.
Metadata curation, Samples and logistics, and Visualisation:
Ms Hannah Lowe 25.
Metadata curation, Sequencing and analysis, and Software and analysis tools:
Dr Ben Temperton 91, Dr Sunando Roy 82, Dr Anna Price 10, Dr Sara Rey 69 and Mr Matthew Wyles 93.
Metadata curation, Sequencing and analysis, and Visualisation:
Stefan Rooke 90 and Dr Sharif Shaaban 68.
Project administration, Samples and logistics, Sequencing and analysis:
Dr Mariateresa de Cesare 98.
Project administration, Samples and logistics, and Software and analysis tools:
Laura Letchford 99.
Project administration, Samples and logistics, and Visualisation:
Miss Siona Silveira 81, Dr Emanuela Pelosi 81 and Dr Eleri Wilson-Davies 81.
Samples and logistics, Sequencing and analysis, and Software and analysis tools:
Dr Myra Hosmillo 24.
Sequencing and analysis, Software and analysis tools, and Visualisation:
Áine O’Toole 90, Dr Andrew R Hesketh 87, Mr Richard Stark 94, Dr Louis du Plessis 23, Dr Chris Ruis 88, Dr Helen Adams 4 and Dr Yann Bourgeois 76.
Funding acquisition, and Leadership and supervision:
Dr Stephen L Michell 91, Prof Dimitris Gramatopoulos 84, 112, Dr Jonathan Edgeworth 12, Prof Judith Breuer
30, 82, Prof John A Todd 98 and Dr Christophe Fraser 5.
Funding acquisition, and Project administration:
Dr David Buck 98 and Michaela John 9.
Leadership and supervision, and Metadata curation:
Dr Gemma L Kay 70.
Leadership and supervision, and Project administration:
Steve Palmer 99, Prof Sharon J Peacock 88, 64 and David Heyburn 69.
Leadership and supervision, and Samples and logistics:
Danni Weldon 99, Dr Esther Robinson 64, 36, Prof Alan McNally 41, 86, Dr Peter Muir 64, Dr Ian B Vipond 64, Dr John BoYes 29, Dr Venkat Sivaprakasam 46, Dr Tranprit Saluja 75, Dr Samir Dervisevic 54 and Dr Emma J Meader 54.
Leadership and supervision, and Sequencing and analysis:
Dr Naomi R Park 99, Karen Oliver 99, Dr Aaron R Jeffries 91, Dr Sascha Ott 94, Dr Ana da Silva Filipe 48, Dr David A Simpson 72 and Dr Chris Williams 69.
Leadership and supervision, and Visualisation:
Dr Jane A H Masoli 73, 91.
Metadata curation, and Samples and logistics:
Dr Bridget A Knight 73, 91, Dr Christopher R Jones 73, 91, Mr Cherian Koshy 1, Miss Amy Ash 1, Dr Anna Casey 71, Dr Andrew Bosworth 64, 36, Dr Liz Ratcliffe 71, Dr Li Xu-McCrae 36, Miss Hannah M Pymont 64, Ms Stephanie Hutchings 64, Dr Lisa Berry 84, Ms Katie Jones 84, Dr Fenella Halstead 46, Mr Thomas Davis 21, Dr Christopher Holmes 16, Prof Miren Iturriza-Gomara 92, Dr Anita O Lucaci 92, Dr Paul Anthony Randell 38, 104, Dr Alison Cox 38, 104, Pinglawathee Madona 38, 104, Dr Kathryn Ann Harris 30, Dr Julianne Rose Brown 30, Dr Tabitha W Mahungu 74, Dr Dianne Irish-Tavares 74, Dr Tanzina Haque 74, Dr Jennifer Hart 74, Mr Eric Witele 74, Mrs Melisa Louise Fenton 75, Mr Steven Liggett 79, Dr Clive Graham 56, Ms Emma Swindells 57, Ms Jennifer Collins 50, Mr Gary Eltringham 50, Ms Sharon Campbell 17, Dr Patrick C McClure 97, Dr Gemma Clark 15, Dr Tim J Sloan 60, Mr Carl Jones 15 and Dr Jessica Lynch 2, 111.
Metadata curation, and Sequencing and analysis:
Dr Ben Warne 8, Steven Leonard 99, Jillian Durham 99, Dr Thomas Williams 90, Dr Sam T Haldenby 92, Dr Nathaniel Storey 30, Dr Nabil-Fareed Alikhan 70, Dr Nadine Holmes 18, Dr Christopher Moore 18, Mr Matthew Carlile 18, Malorie Perry 69, Dr Noel Craine 69, Prof Ronan A Lyons 80, Miss Angela H Beckett 13, Salman Goudarzi 77, Christopher Fearn 77, Kate Cook 77, Hannah Dent 77 and Hannah Paul 77.
Metadata curation, and Software and analysis tools:
Robert Davies 99.
Project administration, and Samples and logistics:
Beth Blane 88, Sophia T Girgis 88, Dr Mathew A Beale 99, Katherine L Bellis 99, 88, Matthew J Dorman 99, Eleanor Drury 99, Leanne Kane 99, Sally Kay 99, Dr Samantha McGuigan 99, Dr Rachel Nelson 99, Liam Prestwood 99, Dr Shavanthi Rajatileka 99, Dr Rahul Batra 12, Dr Rachel J Williams 82, Dr Mark Kristiansen 82, Dr Angie Green 98, Miss Anita Justice 59, Dr Adhyana I.K Mahanama 81, 102 and Dr Buddhini Samaraweera 81, 102.
Project administration, and Sequencing and analysis:
Dr Nazreen F Hadjirin 88 and Dr Joshua Quick 41.
Project administration, and Software and analysis tools:
Mr Radoslaw Poplawski 41.
Samples and logistics, and Sequencing and analysis:
Leanne M Kermack 88, Nicola Reynolds 7, Grant Hall 24, Yasmin Chaudhry 24, Malte L Pinckert 24, Dr Iliana Georgana 24, Dr Robin J Moll 99, Dr Alicia Thornton 66, Dr Richard Myers 66, Dr Joanne Stockton 41, Miss Charlotte A Williams 82, Dr Wen C Yew 58, Alexander J Trotter 70, Miss Amy Trebes 98, Mr George MacIntyre-Cockett 98, Alec Birchley 69, Alexander Adams 69, Amy Plimmer 69, Bree Gatica-Wilcox 69, Dr Caoimhe McKerr 69, Ember Hilvers 69, Hannah Jones 69, Dr Hibo Asad 69, Jason Coombes 69, Johnathan M Evans 69, Laia Fina 69, Lauren Gilbert 69, Lee Graham 69, Michelle Cronin 69, Sara
Kumziene-SummerhaYes 69, Sarah Taylor 69, Sophie Jones 69, Miss Danielle C Groves 93, Mrs Peijun Zhang 93, Miss Marta Gallis 93 and Miss Stavroula F Louka 93.
Samples and logistics, and Software and analysis tools:
Dr Igor Starinskij 48.
Sequencing and analysis, and Software and analysis tools:
Dr Chris J Illingworth 47, Dr Chris Jackson 47, Ms Marina Gourtovaia 99, Gerry Tonkin-Hill 99, Kevin Lewis 99, Dr Jaime M Tovar-Corona 99, Dr Keith James 99, Dr Laura Baxter 94, Dr Mohammad T. Alam 94, Dr Richard J Orton 48, Dr Joseph Hughes 48, Dr Sreenu Vattipally 48, Dr Manon Ragonnet-Cronin 39, Dr Fabricia F. Nascimento 39, Mr David Jorgensen 39, Ms Olivia Boyd 39, Ms Lily Geidelberg 39, Dr Alex E Zarebski 23, Dr Jayna Raghwani 23, Dr Moritz UG Kraemer 23, Joel Southgate 10, 69, Dr Benjamin B Lindsey 93 and Mr Timothy M Freeman 93.
Software and analysis tools, and Visualisation:
Jon-Paul Keatley 99, Dr Joshua B Singer 48, Leonardo de Oliveira Martins 70, Dr Corin A Yeats 14, Dr Khalil Abudahab 14, 114, Mr Ben EW Taylor 14, 114 and Mirko Menegazzo 14.
Leadership and supervision:
Prof John Danesh 99, Wendy Hogsden 46, Dr Sahar Eldirdiri 21, Mrs Anita Kenyon 21, Dr Jenifer Mason 43, Mr Trevor I Robinson 43, Prof Alison Holmes 38, 103, Dr James Price 38, 103, Prof John A Hartley 82, Dr Tanya Curran 3, Dr Alison E Mather 70, Dr Giri Shankar 69, Dr Rachel Jones 69, Dr Robin Howe 69 and Dr Sian Morgan 9.
Metadata curation:
Dr Elizabeth Wastenge 53, Dr Michael R Chapman 34, 88, 99, Mr Siddharth Mookerjee 38, 103, Dr Rachael Stanley 54, Mrs Wendy Smith 15, Prof Timothy Peto 59, Dr David Eyre 59, Dr Derrick Crook 59, Dr Gabrielle Vernet 33, Dr Christine Kitchen 10, Huw Gulliver 10, Dr Ian Merrick 10, Prof Martyn Guest 10, Robert Munn 10, Dr Declan T Bradley 63, 72, and Dr Tim Wyatt 63.
Project administration:
Dr Charlotte Beaver 99, Luke Foulser 99, Sophie Palmer 88, Carol M Churcher 88, Ellena Brooks 88, Kim S Smith 88, Dr Katerina Galai 88, Georgina M McManus 88, Dr Frances Bolt 38, 103, Dr Francesc Coll 19, Lizzie Meadows 70, Dr Stephen W Attwood 23, Dr Alisha Davies 69, Elen De Lacy 69, Fatima Downing 69, Sue Edwards 69, Dr Garry P Scarlett 76, Mrs Sarah Jeremiah 83 and Dr Nikki Smith 93.
Samples and logistics:
Danielle Leek 88, Sushmita Sridhar 88, 99, Sally Forrest 88, Claire Cormie 88, Harmeet K Gill 88, Joana Dias 88, Ellen E Higginson 88, Mailis Maes 88, Jamie Young 88, Michelle Wantoch 7, Sanger Covid Team (www.sanger.ac.uk/covid-team) 99, Dorota Jamrozy 99, Stephanie Lo 99, Dr Minal Patel 99, Verity Hill 90, Ms Claire M Bewshea 91, Prof Sian Ellard 73, 91, Dr Cressida Auckland 73, Dr Ian Harrison 66, Dr Chloe Bishop 66, Dr Vicki Chalker 66, Dr Alex Richter 85, Dr Andrew Beggs 85, Dr Angus Best 86, Dr Benita Percival 86, Dr Jeremy Mirza 86, Dr Oliver Megram 86, Dr Megan Mayhew 86, Dr Liam Crawford 86, Dr Fiona Ashcroft 86, Dr Emma Moles-Garcia 86, Dr Nicola Cumley 86, Mr Richard Hopes 64, Dr Patawee Asamaphan 48, Mr Marc O Niebel 48, Prof Rory N Gunson 100, Dr Amanda Bradley 52, Dr Alasdair Maclean 52, Dr Guy Mollett 52, Dr Rachel Blacow 52, Mr Paul Bird 16, Mr Thomas Helmer 16, Miss Karlie Fallon 16, Dr Julian Tang 16, Dr Antony D Hale 49, Dr Louissa R Macfarlane-Smith 49, Katherine L Harper 49, Miss Holli Carden 49, Dr Nicholas W Machin 45, 64, Ms Kathryn A Jackson 92, Dr Shazaad S Y Ahmad 45, 64, Dr Ryan P George 45, Dr Lance Turtle 92, Mrs Elaine O’Toole 43, Mrs Joanne Watts 43, Mrs Cassie Breen 43, Mrs Angela Cowell 43, Ms Adela Alcolea-Medina 32, 96, Ms Themoula Charalampous 12, 42, Amita Patel 11, Dr Lisa J Levett 35, Dr Judith Heaney 35, Dr Aileen Rowan 39, Prof Graham P Taylor 39, Dr Divya Shah 30, Miss Laura Atkinson 30, Mr Jack CD Lee 30, Mr Adam P Westhorpe 82, Dr Riaz Jannoo 82, Dr Helen L Lowe 82, Miss Angeliki Karamani 82, Miss Leah Ensell 82, Mrs Wendy Chatterton 35, Miss Monika Pusok 35, Mrs Ashok Dadrah 75, Miss Amanda Symmonds 75, Dr Graciela Sluga 44, Dr Zoltan Molnar 72, Mr Paul Baker 79, Prof Stephen Bonner 79, Ms Sarah Essex 79, Dr Edward Barton 56, Ms Debra Padgett 56, Ms Garren Scott 56, Ms Jane Greenaway 57, Dr Brendan AI Payne 50, Dr Shirelle Burton-Fanning 50, Dr Sheila Waugh 50, Dr Veena Raviprakash 17, Ms Nicola Sheriff 17, Ms Victoria Blakey 17, ms Lesley-Anne Williams 17, Dr Jonathan Moore 27, Ms Susanne Stonehouse 27, Dr Louise Smith 55, Dr Rose K Davidson 89, Dr Luke Bedford 26, Dr Lindsay Coupland 54, Ms Victoria Wright 18, Dr Joseph G Chappell 97, Dr Theocharis Tsoleridis 97, Prof Jonathan Ball 97, Mrs Manjinder Khakh 15, Dr Vicki M Fleming 15, Dr Michelle M Lister 15, Dr Hannah C Howson-Wells 15, Dr Louise Berry 15, Dr Tim Boswell 15, Dr Amelia Joseph 15, Dr Iona Willingham 15, Dr Nichola Duckworth 60, Dr Sarah Walsh 60, Dr Emma Wise 2, 111, Dr Nathan Moore 2, 111, Miss Matilde Mori 2, 108, 111, Dr Nick Cortes 2, 111, Dr Stephen Kidd 2, 111, Dr Rebecca Williams 33, Laura Gifford 69, Miss Kelly Bicknell 61, Dr Sarah Wyllie 61, Miss Allyson Lloyd 61, Mr Robert Impey 61, Ms Cassandra S Malone 6, Mr Benjamin J Cogger 6, Nick Levene 62, Lynn Monaghan 62, Dr Alexander J Keeley 93, Dr David G Partridge 78, 93, Dr Mohammad Raza 78, 93, Dr Cariad Evans 78, 93 and Dr Kate Johnson 78, 93.
Sequencing and analysis:
Emma Betteridge 99, Ben W Farr 99, Scott Goodwin 99, Dr Michael A Quail 99, Carol Scott 99, Lesley Shirley 99, Scott AJ Thurston 99, Diana Rajan 99, Dr Iraad F Bronner 99, Louise Aigrain 99, Dr Nicholas M Redshaw 99, Dr Stefanie V Lensing 99, Shane McCarthy 99, Alex Makunin 99, Dr Carlos E Balcazar 90, Dr Michael D Gallagher 90, Dr Kathleen A Williamson 90, Thomas D Stanton 90, Ms Michelle L Michelsen 91, Ms Joanna Warwick-Dugdale 91, Dr Robin Manley 91, Ms Audrey Farbos 91, Dr James W Harrison 91, Dr Christine M Sambles 91, Dr David J Studholme 91, Dr Angie Lackenby 66, Dr Tamyo Mbisa 66, Dr Steven Platt 66, Mr Shahjahan Miah 66, Dr David Bibby 66, Dr Carmen Manso 66, Dr Jonathan Hubb 66, Dr Gavin Dabrera 66, Dr Mary Ramsay 66, Dr Daniel Bradshaw 66, Dr Ulf Schaefer 66, Dr Natalie Groves 66, Dr Eileen Gallagher 66, Dr David Lee 66, Dr David Williams 66, Dr Nicholas Ellaby 66, Hassan Hartman 66, Nikos Manesis 66, Vineet Patel 66, Juan Ledesma 67, Ms Katherine A Twohig 67, Dr Elias Allara 64, 88, Ms Clare Pearson 64, 88, Mr Jeffrey K. J. Cheng 94, Dr Hannah E. Bridgewater 94, Ms Lucy R. Frost 94, Ms Grace Taylor-Joyce 94, Dr Paul E Brown 94, Dr Lily Tong 48, Ms Alice Broos 48, Mr Daniel Mair 48, Mrs Jenna Nichols 48, Dr Stephen N Carmichael 48, Dr Katherine L Smollett 40, Dr Kyriaki Nomikou 48, Dr Elihu Aranday-Cortes 48, Ms Natasha Johnson 48, Dr Seema Nickbakhsh 48, 68, Dr Edith E Vamos 92, Dr Margaret Hughes 92, Dr Lucille Rainbow 92, Mr Richard Eccles 92, Ms Charlotte Nelson 92, Dr Mark Whitehead 92, Dr Richard Gregory 92, Mr Matthew Gemmell 92, Ms Claudia Wierzbicki 92, Ms Hermione J Webster 92, Ms Chloe L Fisher 28, Mr Adrian W Signell 20, Dr Gilberto Betancor 20, Mr Harry D Wilson 20, Dr Gaia Nebbia 12, Dr Flavia Flaviani 31, Mr Alberto C Cerda 96, Ms Tammy V Merrill 96, Rebekah E Wilson 96, Mr Marius Cotic 82, Miss Nadua Bayzid 82, Dr Thomas Thompson 72, Dr Erwan Acheson 72, Prof Steven Rushton 51, Prof Sarah O’Brien 51, David J Baker 70, Steven Rudder 70, Alp Aydin 70, Dr Fei Sang 18, Dr Johnny Debebe 18, Dr Sarah Francois 23, Dr Tetyana I Vasylyeva 23, Dr Marina Escalera Zamudio 23, Mr Bernardo Gutierrez 23, Dr Angela Marchbank 10, Joshua Maksimovic 9, Karla Spellman 9, Kathryn McCluggage 9, Dr Mari Morgan 69, Robert Beer 9, Safiah Afifi 9, Trudy Workman 10, William Fuller 10, Catherine Bresner 10, Dr Adrienn Angyal 93, Dr Luke R Green 93, Dr Paul J Parsons 93, Miss Rachel M Tucker 93, Dr Rebecca Brown 93 and Mr Max Whiteley 93.
Software and analysis tools:
James Bonfield 99, Dr Christoph Puethe 99, Mr Andrew Whitwham 99, Jennifier Liddle 99, Dr Will Rowe 41, Dr Igor Siveroni 39, Dr Thanh Le-Viet 70 and Amy Gaskin 69.
Visualisation:
Dr Rob Johnson 39.
1 Barking, Havering and Redbridge University Hospitals NHS Trust, 2 Basingstoke Hospital, 3 Belfast Health & Social Care Trust, 4 Betsi Cadwaladr University Health Board, 5 Big Data Institute, Nuffield Department of Medicine, University of Oxford, 6 Brighton and Sussex University Hospitals NHS Trust, 7 Cambridge Stem Cell Institute, University of Cambridge, 8 Cambridge University Hospitals NHS Foundation Trust, 9 Cardiff and Vale University Health Board, 10 Cardiff University, 11 Centre for Clinical Infection & Diagnostics Research, St. Thomas’ Hospital and Kings College London, 12 Centre for Clinical Infection and Diagnostics Research, Department of Infectious Diseases, Guy’s and St Thomas’ NHS Foundation Trust, 13 Centre for Enzyme Innovation, University of Portsmouth (PORT), 14 Centre for Genomic Pathogen Surveillance, University of Oxford, 15 Clinical Microbiology Department, Queens Medical Centre, 16 Clinical Microbiology, University Hospitals of Leicester NHS Trust, 17 County Durham and Darlington NHS Foundation Trust, 18 Deep Seq, School of Life Sciences, Queens Medical Centre, University of Nottingham, 19 Department of Infection Biology, Faculty of Infectious & Tropical Diseases, London School of Hygiene & Tropical Medicine, 20 Department of Infectious Diseases, King’s College London, 21 Department of Microbiology, Kettering General Hospital, 22 Departments of Infectious Diseases and Microbiology, Cambridge University Hospitals NHS Foundation Trust; Cambridge, UK, 23 Department of Zoology, University of Oxford, 24 Division of Virology, Department of Pathology, University of Cambridge, 25 East Kent Hospitals University NHS Foundation Trust, 26 East Suffolk and North Essex NHS Foundation Trust, 27 Gateshead Health NHS Foundation Trust, 28 Genomics Innovation Unit, Guy’s and St. Thomas’ NHS Foundation Trust, 29 Gloucestershire Hospitals NHS Foundation Trust, 30 Great Ormond Street Hospital for Children NHS Foundation Trust, 31 Guy’s and St. Thomas’ BRC, 32 Guy’s and St. Thomas’ Hospitals, 33 Hampshire Hospitals NHS Foundation Trust, 34 Health Data Research UK Cambridge, 35 Health Services Laboratories, 36 Heartlands Hospital, Birmingham, 37 Hub for Biotechnology in the Built Environment, Northumbria University, 38 Imperial College Hospitals NHS Trust, 39 Imperial College London, 40 Institute of Biodiversity, Animal Health & Comparative Medicine, 41 Institute of Microbiology and Infection, University of Birmingham, 42 King’s College London, 43 Liverpool Clinical Laboratories, 44 Maidstone and Tunbridge Wells NHS Trust, 45 Manchester University NHS Foundation Trust, 46 Microbiology Department, Wye Valley NHS Trust, Hereford, 47 MRC Biostatistics Unit, University of Cambridge, 48 MRC-University of Glasgow Centre for Virus Research, 49 National Infection Service, PHE and Leeds Teaching Hospitals Trust, 50 Newcastle Hospitals NHS Foundation Trust, 51 Newcastle University, 52 NHS Greater Glasgow and Clyde, 53 NHS Lothian, 54 Norfolk and Norwich University Hospital, 55 Norfolk County Council, 56 North Cumbria Integrated Care NHS Foundation Trust, 57 North Tees and Hartlepool NHS Foundation Trust, 58 Northumbria University, 59 Oxford University Hospitals NHS Foundation Trust, 60 PathLinks, Northern Lincolnshire & Goole NHS Foundation Trust, 61 Portsmouth Hospitals University NHS Trust, 62 Princess Alexandra Hospital Microbiology Dept., 63 Public Health Agency, 64 Public Health England, 65 Public Health England, Clinical Microbiology and Public Health Laboratory, Cambridge, UK, 66 Public Health England, Colindale, 67 Public Health England, Colindale, 68 Public Health Scotland, 69 Public Health Wales NHS Trust, 70 Quadram Institute Bioscience, 71 Queen Elizabeth Hospital, 72 Queen’s University Belfast, 73 Royal Devon and Exeter NHS Foundation Trust, 74 Royal Free NHS Trust, 75 Sandwell and West Birmingham NHS Trust, 76 School of Biological Sciences, University of Portsmouth (PORT), 77 School of Pharmacy and Biomedical Sciences, University of Portsmouth (PORT), 78 Sheffield Teaching Hospitals, 79 South Tees Hospitals NHS Foundation Trust, 80 Swansea University, 81 University Hospitals Southampton NHS Foundation Trust, 82 University College London, 83 University Hospital Southampton NHS Foundation Trust, 84 University Hospitals Coventry and Warwickshire, 85 University of Birmingham, 86 University of Birmingham Turnkey Laboratory, 87 University of Brighton, 88 University of Cambridge, 89 University of East Anglia, 90 University of Edinburgh, 91 University of Exeter, 92 University of Liverpool, 93 University of Sheffield, 94 University of Warwick, 95 University of Cambridge, 96 Viapath, Guy’s and St Thomas’ NHS Foundation Trust, and King’s College Hospital NHS Foundation Trust, 97 Virology, School of Life Sciences, Queens Medical Centre, University of Nottingham, 98 Wellcome Centre for Human Genetics, Nuffield Department of Medicine, University of Oxford, 99 Wellcome Sanger Institute, 100 West of Scotland Specialist Virology Centre, NHS Greater Glasgow and Clyde, 101 Department of Medicine, University of Cambridge, 102 Ministry of Health, Sri Lanka, 103 NIHR Health Protection Research Unit in HCAI and AMR, Imperial College London, 104 North West London Pathology, 105 NU-OMICS, Northumbria University, 106 University of Kent, 107 University of Oxford, 108 University of Southampton, 109 University of Southampton School of Health Sciences, 110 University of Southampton School of Medicine, 111 University of Surrey, 112 Warwick Medical School and Institute of Precision Diagnostics, Pathology, UHCW NHS Trust, 113 Wellcome Africa Health Research Institute Durban and 114 Wellcome Genome Campus.
Footnotes
Version 1 had a figure 3 missing which has been added and in SI section a link to code and data is inserted.
↵1 We calculated this as the posterior mean and 2.5-97.5 quantiles of the set of 42×6 posterior medians of the distributions (R_{S-} - R_{S+}), one per STP-week
References




