
Abstract
SARS-CoV-2 has infected millions of people globally. Virus infection requires the receptor-binding domain (RBD) of the spike protein. Although studies have demonstrated anti-spike and - RBD antibodies to be protective in animal models and convalescent plasma as a promising therapeutic option, little is known about immunoglobulin (Ig) isotypes capable of blocking infection. Here, we studied spike- and RBD-specific Ig isotypes in convalescent and acute plasma/sera. We also determined virus neutralization activities in plasma/sera, and purified Ig fractions. Spike- and RBD-specific IgM, IgG1, and IgA1 were produced by all or nearly all subjects at variable levels and detected early after infection. All samples also displayed neutralizing activity. Regression analyses revealed that IgM and IgG1 contributed most to neutralization, consistent with IgM and IgG fractions’ neutralization potency. However, IgA also exhibited neutralizing activity at a lower potency. Together, IgG, IgM and IgA are critical components of convalescent plasma used for COVID-19 treatment.
- SARS-CoV-2
- COVID-19
- antibody isotypes
- neutralization
- convalescent plasma
Background
In December 2019, the first patients with coronavirus disease 2019 (COVID-19), caused by severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), were identified in Wuhan’s city of Hubei Province, China1. Since then, the epidemic has rapidly spread to most regions of the world, infecting millions of people2. Effective therapeutics and vaccines against SARS-CoV-2 are urgently needed. Convalescent plasma transfusion showed promising results in patients with severe to life-threatening COVID-193–7 and clinical trials to evaluate the efficacy of convalescent plasma treatment for ambulatory and hospitalized COVID-19 patients are underway8–11. To this end, more information about the Ig isotypes present in the plasma of COVID-19 convalescent individuals and their antiviral activities are needed. It is also unclear which of the immunoglobulin (Ig) isotypes present in convalescent plasma are protective. The data would likewise inform vaccine development, as more than 100 vaccine candidates are in different stages of preclinical development, and many are now in phase 2 and 3 clinical trials12. Although using different strategies13, most vaccines are based on the SARS-CoV-2 spike protein14,15, which is a membrane-anchored protein present with two others (membrane and envelope proteins) on the virus envelope surface and contains the receptor-binding domain (RBD) required for binding to and entry into the cells16–22. These vaccines aim to protect by inducing neutralizing antibodies (Abs) capable of blocking viral infection.
Monomeric IgG constitutes approximately 75% of the Abs found in serum and exists as four subtypes: IgG1 (∼66% of IgG), IgG2 (∼23% of IgG), IgG3 (∼7% of IgG) and IgG4 (∼4% of IgG)23,24. IgM Abs represent 10% of total serum Abs and are the first to arise in response to new antigens24,25. Although IgM Abs do not undergo extensive somatic hypermutation to increase their affinity as do IgG and IgA Abs, their higher valency due to oligomerization enhances their avidity and potency against pathogens24,26,27. IgA Abs exist as two subtypes: IgA1 and IgA2, representing 15% of total serum Abs. IgA1 is the primary IgA subtype in serum while IgA2 predominates in mucosal secretions24. These two IgA subtypes are dimeric in the mucosa, but in the circulation, they are monomeric.
SARS-CoV-2 spike-, RBD- and nucleocapsid-specific serum/plasma Abs of IgM, IgG, and IgA isotypes are found in most COVID-19 patients28–35, with Ab neutralizing activities reported developing within the first two weeks of infection and decline over time31,33,36–38. However, the neutralizing titers appear to vary greatly31,33,36–38, and they correlate with Ab binding levels against RBD, spike, and/or nucleocapsid, and with age, duration of symptoms, and symptom severity31,33,37. Several RBD-specific monoclonal Abs of IgG isotype with potent antiviral activities have been generated from individuals with high neutralization titers, and these confer protection in animal models31,36,39,40. Moreover, a monoclonal Ab of IgA isotype capable of recognizing both the SARS-CoV-1 and SARS-CoV-2 spike proteins and blocking ACE2 receptor binding was recently described41. However, no direct evidence is available regarding the neutralizing capacity of plasma IgM and IgA Abs from COVID-19 patients.
Studies on other respiratory viruses such as influenza show that, in addition to IgG, IgA could also mediate virus neutralization, and their relative contribution depends on the physiologic compartment in which they are found, with IgA contributing to the protection of mostly the upper respiratory tract while IgG was protective of the lower respiratory tract42–45. Of note, an anti-hemagglutinin monoclonal polymeric IgA has been demonstrated to mediate more potent antiviral activities against influenza when compared to a monoclonal IgG against the same epitope46. Interestingly, an IgM Ab with potent antiviral activities targeting influenza B’s receptor binding site has also been described47. In addition, respiratory syncytial virus (RSV)-specific mucosal IgA neutralizing Abs are a better correlate of protection than RSV-specific serum IgG neutralizing Abs48. In the case of SARS-CoV-1, high titers of mucosal IgA in the lungs correlated with reduced pathology upon viral challenge in animal models49. Whether IgA in the blood and in the respiratory tract’s mucosa offers protection against SARS-CoV-2 infection or COVID-19 disease remains an open question. Moreover, few data are available regarding the contribution of IgM to neutralization and protection against viruses, including SARS-CoV-2. Of note, in terminally ill COVID-19 patients, systemic SARS-CoV-2 infection affecting multiple organs including the heart, kidneys, and brain, is evident from autopsy studies50–52. Thus, the capacity of plasma Ig to suppress virus spread to these organs is critical for effective convalescent plasma therapy against severe COVID-19 disease.
We recently described a multiplex bead Ab binding assay using the Luminex technology to detect total Ig against spike and RBD53. Based on this assay, we characterized the Ig isotype profiles against spike and RBD in the plasma and serum from acutely infected or convalescent individuals using this Luminex assay which detects antigen-specific IgM, IgG1-4, IgA1, and IgA2. Using a pseudovirus assay54, we also measured neutralizing activities in plasma, serum, and Ig isotype plasma fractions to determine the neutralizing capacity of IgM, IgA, and IgG. The data indicated a high prevalence of spike- and RBD-specific IgM and IgA Abs, similar to that of IgG1, in plasma and serum from COVID-19 patients, and their contributions to virus neutralizing activities. By testing purified IgG, IgM and IgA Abs from plasma of convalescent COVID-19 subjects, this study presents the first direct evidence that plasma IgG, IgM, and IgA all contribute to SARS-CoV-2 neutralization.
Results
Levels of Ig isotypes against the SARS-CoV-2 spike and RBD vary in convalescent Individuals
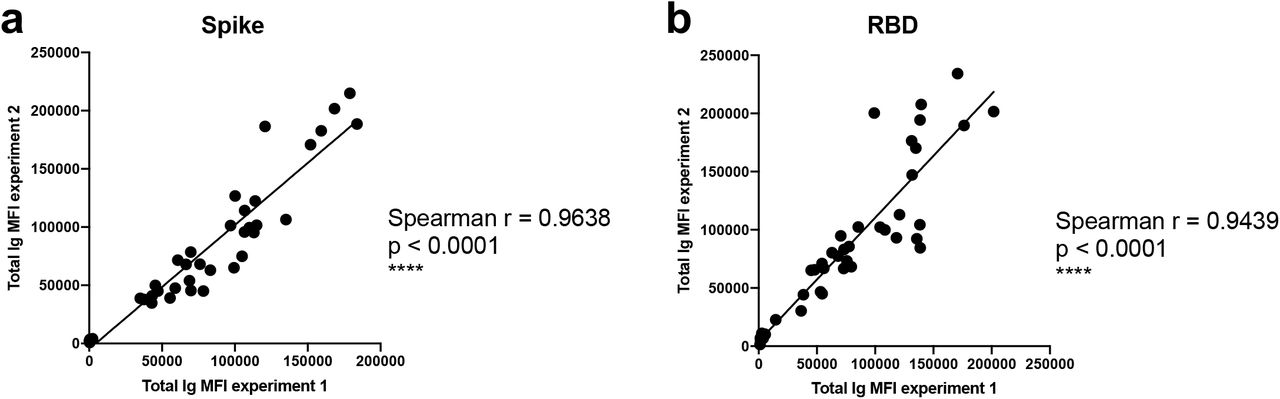
A total of 29 serum (P#5-8) and plasma (TF#1-25) specimens from COVID-19-convalescent individuals was tested. TF#1-25 were collected between March 26 and April 7, 2020, about 4-8 weeks after the initial outbreak in North American, and used for transfusion into hospitalized COVID-19 patients3. Ten plasma from COVID-negative contemporaneous blood bank donors (N#4-13) were included for comparison. Sera or plasma from 12 uninfected individuals banked prior to the COVID-19 outbreak (N#1-3 and N#14-22) were used to establish background values. The specimens were initially titrated for total Ig against spike and RBD (Fig. 1). All 29 COVID-19 positive specimens exhibited titration curves of total Ig Abs against spike, while none of the negative controls did. Similar results were observed with RBD, except that one contemporaneous COVID-19-negative sample had low levels of RBD-specific Ig (N#10). Overall the background MFI values were higher for RBD than spike. To assess the reproducibility of the assay, the samples were tested in at least two separate experiments run on different days, and a strong correlation was observed between the MFI values from these independent experiments (Supplementary Fig. 1). The areas under the curves (AUCs) highly correlated with the MFI values from specimens diluted 1:200 (p < 0.0001; Supplementary Fig. 2); consequently, all samples were tested for isotyping at this dilution. At the 1:200 dilution we were able to discern a diverse range of Ig isotype levels among individual samples (Fig. 2). To evaluate for the presence of spike-specific and RBD-specific total Ig, IgM, IgG1, IgG2, IgG3, IgG4, IgA1 and IgA2, the specificity and strength of the secondary Abs used to detect the different isotypes were first validated with Luminex beads coated with myeloma proteins of known Ig isotypes (IgG1, IgG2, IgG3, IgG4, IgA1, IgA2, and IgM). All eight secondary Abs were able to detect their specific Ig isotypes with MFI values reaching > 60,000 (Supplementary Fig. 3).
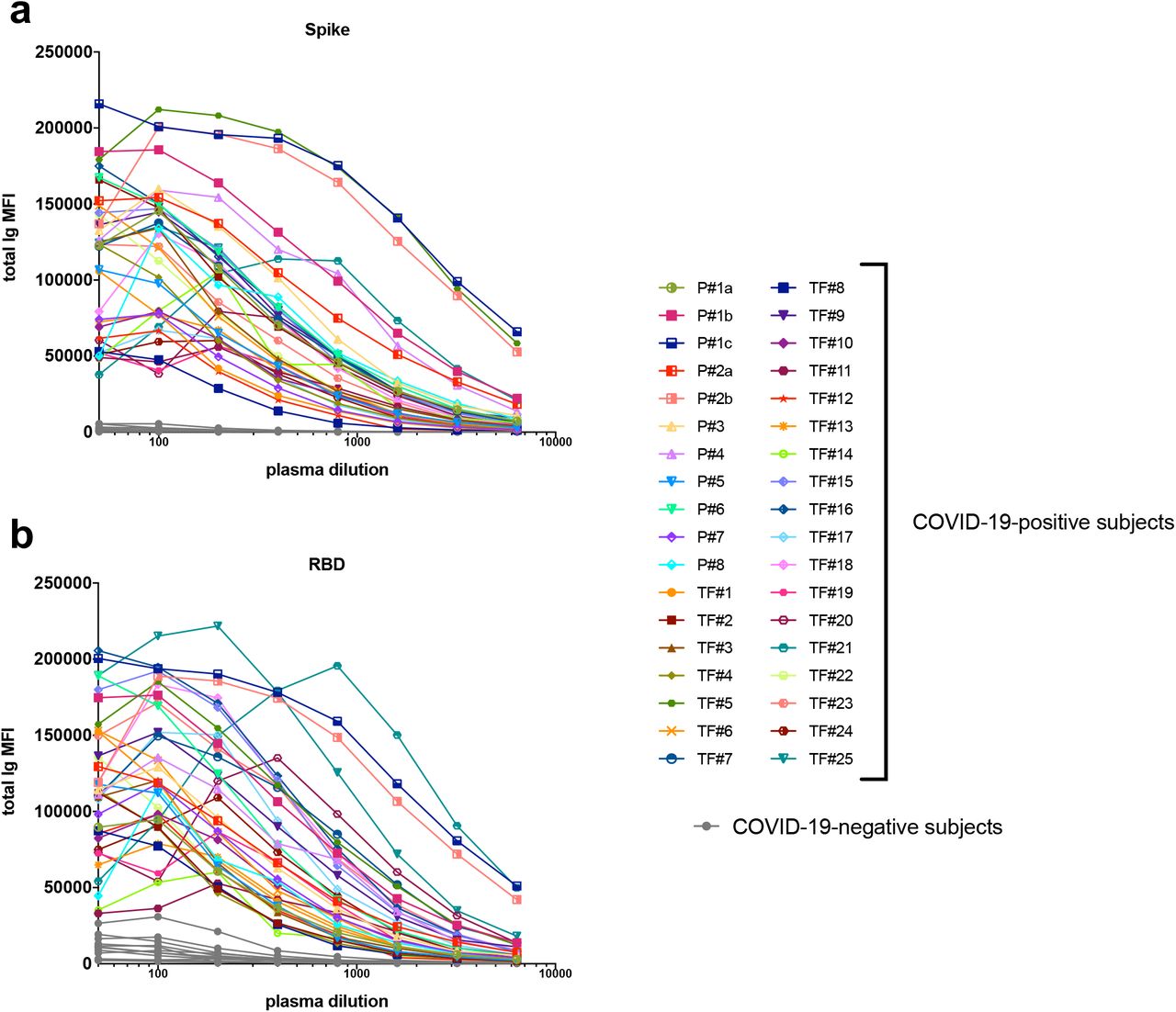
Titration of (a) spike-specific or (b) RBD-specific total Ig from 29 COVID-19-convalescent individuals, two acute COVID-19 patients with longitudinal samples, and 13 COVID-19-negative individuals. Specimens were diluted at 2-fold dilutions from 1:50 to 1:6,400.
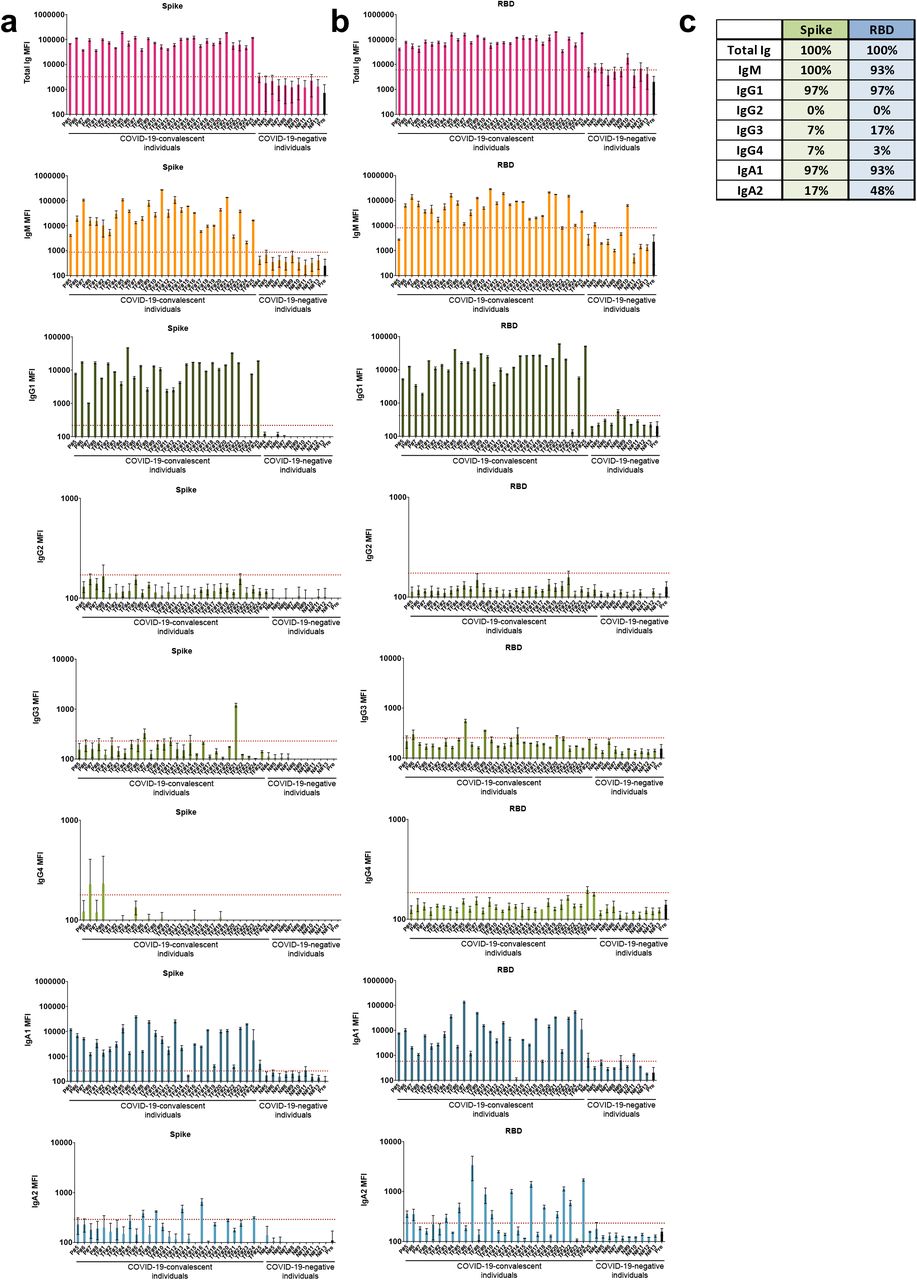
Detection of total Ig, IgM, IgG1, IgG2, IgG3, IgG4, IgA1 and IgA2 against (a) spike and (b) RBD in specimens from 29 COVID-19-convalescent individuals, 13 COVID-19-negative contemporary samples, and pre-pandemic controls. The samples were tested at a dilution of 1:200 and data are shown as mean MFI + standard deviation (SD) of duplicate measurements from at least two independent experiments. The pre-pandemic controls are shown as mean MFI + SD of 12 samples (Pre, black bar). The horizontal red dotted line represents the cut-off value determined as the mean + 3 SD of 12 pre-pandemic samples for each of the isotypes. (c) Percentages of responders above the cut-off for each spike-or RBD-specific Ig isotype.
All 29 convalescent individuals had anti-spike and anti-RBD total Ig (Fig. 2), but the Ig levels were highly variable, with MFI values ranging from 36,083 to 190,150. In addition, all 29 convalescent individuals also displayed IgM Abs against spike at varying levels, and 93% were positive for anti-RBD IgM when evaluated using cut-off values calculated as mean + 3 standard deviation (SD) of the 12 pre-pandemic samples (Fig 2b, c). An IgG1 response was detected against both spike and RBD in 97% of the convalescent subjects, with MFI values that ranged from 1,013 to 59,880. In contrast, IgG2, IgG3, and IgG4 Abs against spike and RBD were detected in only a small fraction of the subjects, and the levels were very low (MFI values < 1,300) (Fig. 2). Surprisingly, almost all individuals produced IgA1 Abs against spike (97%) and RBD (93%), while 17% exhibited IgA2 against spike, and 48% exhibited IgA2 against RBD (Fig. 2). Low levels, slightly above cut-off, of spike- and RBD-binding total Ig, IgM, and IgG1, and IgA1 were also detected sporadically in contemporaneous COVID-19 samples, such as N#8, N#10, and N#11. The responses against spike and RBD were highly correlated for every isotype (Supplementary Fig. 4). Overall, these data demonstrate that IgM, IgG1, and IgA1 Abs were induced against spike and RBD in all or almost all COVID-19 convalescent individuals (Fig. 2). The levels, however, were highly variable among individuals. No statistical significance in the levels of total Ig, IgM, IgG1, and IgA1 was observed between female and male individuals (Supplementary Fig. 5).
In Fig. 3, regression analyses to assess the impact of individual isotypes on the total Ig binding showed that IgG1 had the highest r2 values (0.83 and 0.70 for spike- and RBD-binding IgG1, respectively) with p <0.0001, indicating that IgG1 is the major isotype induced by SARS-CoV-2 infection against spike and RBD (Fig. 3a,b). IgG2 Abs against RBD had an r2 value of 0.55 with p < 0.0001, but IgG2 levels were very low. For all other isotypes, including IgM, the r2 values were less than 0.40 (Fig. 3c). Thus, despite the presence of many isotypes in sera and plasma, as expected, the major isotype is IgG1.

Simple linear regression of (a) spike-specific or (b) RBD-specific total Ig levels versus IgM, IgG1 or IgG2 levels or versus (c) spike-specific and RBD-specific IgG3, IgG4, IgA1, and IgA2 levels from the 29 COVID-19-convalescent individuals from Fig. 1. The dash lines represent 95% confidence intervals.
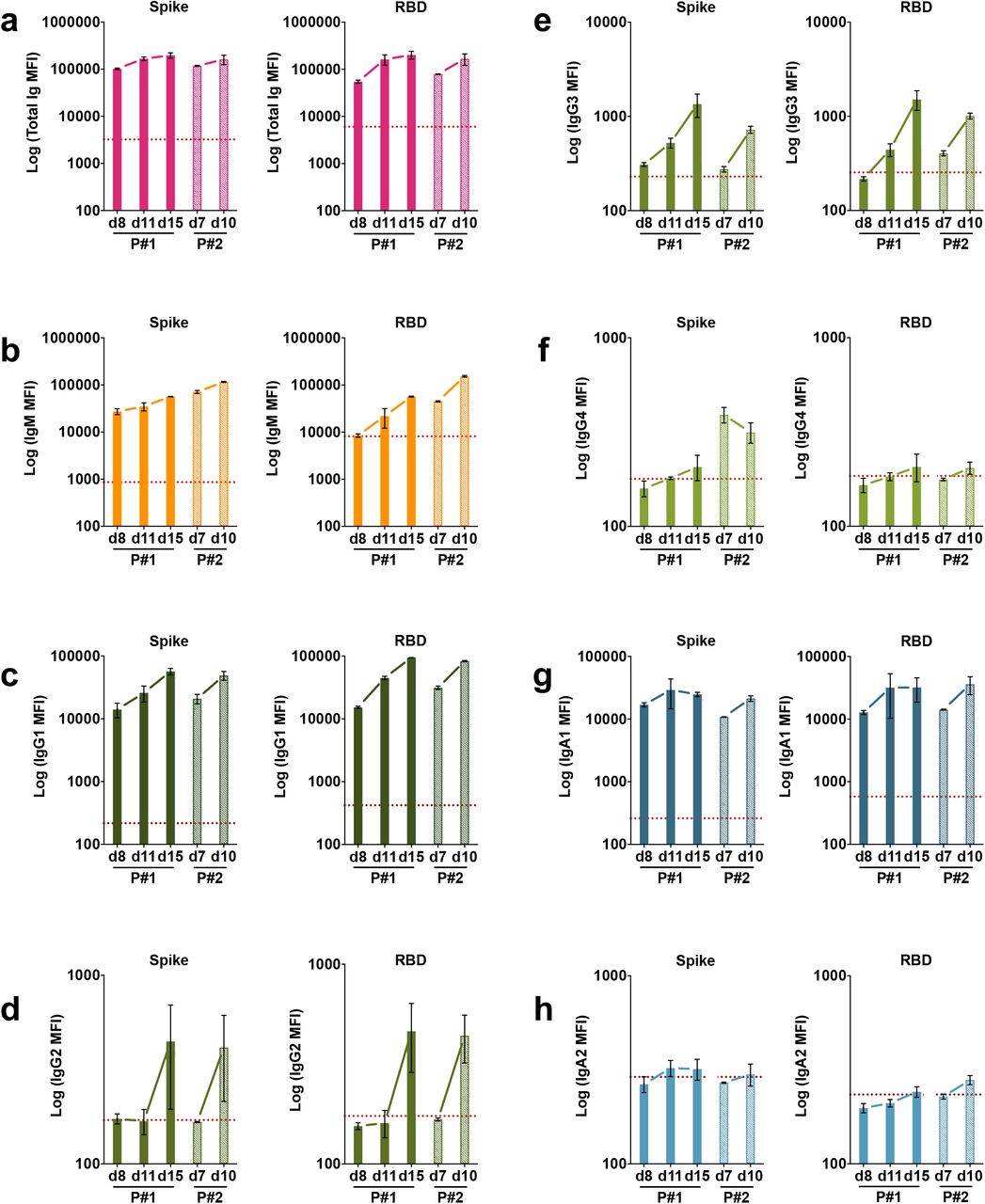
Specimens from two patients (P#1 and P#2) were drawn during the acute phase of the infection. Serial specimens from these patients were tested to determine the isotypes of Abs present early in infection. The earliest samples from both patients, drawn at 7 or 8 days after symptom onset were already positive for total Ig, IgG1, IgA1and IgM Abs against spike (Supplementary Fig. 6a) and RBD (Supplementary Fig. 6b), and these levels increased over the following three to seven days. On the contrary, IgA2 Ab levels were near or below background on days 7-8 and remained unchanged over two weeks post-onset. IgG4 Abs also remained low or near background, whereas IgG2 and IgG3 Abs increased slightly to above background after 10-15 days.
Neutralizing activities are detected in all convalescent COVID-19 individuals
We subsequently tested the ability of samples from convalescent subjects to neutralize a VSVΔG pseudovirus bearing the SARS-CoV-2 spike protein (COV2pp). This pseudovirus assay demonstrated a strong positive correlation with neutralization of the authentic SARS-CoV-2 virus (p = 0.82 and p < 0.0001 for IC50 correlation)54. The results, shown in Fig. 4, demonstrate the percentages of COV2pp neutralizing activity in serum or plasma specimens from 28 COVID-19-convalescent individuals and 11 COVID-19-negative individuals over a range of seven serial four-fold dilutions. A soluble recombinant RBD (sRBD) protein capable of blocking virus infection was tested in parallel as a positive control.

Neutralization of COV2pp with (a,b,c) WT or (d,e) D614G mutated spike proteins by samples from (a,d) 28 COVID-19-convalescent individuals and (b,e) 11 COVID-19-negative individuals, compared to (c) a recombinant soluble RBD (sRBD) control. Each plasma or sera specimen was tested at 4-fold dilutions from 1:10 to 1:40,960, and sRBD was tested at 4-fold dilutions from 100 to 0.02 µg/mL. The data are shown as mean percentage of neutralization + SD of triplicate. The extrapolated titration curves were generated using a nonlinear regression model in GraphPad Prism (Inhibitor versus response – variable slope (four parameters), least squares regression). The dotted horizontal lines highlight 50% neutralization. (f) Spearman correlation between the IC90 titers against COV2pp WT versus D614G.
All specimens from COVID-19-convalescent individuals were able to neutralize the virus at levels above 50% (Fig. 4a). For 26 of 28 specimens, neutralization reached more than 90% (Fig. 4a). The sample with the lowest titer (reciprocal IC50 titer = 37) reached a neutralization plateau of only ∼60%. Of note, one sample (TF#11) demonstrated highly potent neutralization with a reciprocal IC50 titer > 40,960, and neutralization was still 75% at the highest dilution tested. None of the samples from COVID-19-negative individuals reached 50% neutralization (Fig. 4b), while the sRBD positive control demonstrated potent neutralization with an IC50 of 0.06 µg/mL (Fig. 4c), similar to that recently reported54.
The samples were also tested for neutralization against a COV2pp bearing spike with a D614 mutation (D614G mutant), as the D614G variant has become the most prevalent circulating strain in the global pandemic55. Similar to the WT COV2pp, all COVID-19-convalescent samples had neutralizing activity reaching >50% (Fig. 4d), while none of the negative samples did (Fig. 4e). The IC90 titers against WT and D614 mutant differed on average by only 1.7-fold and correlated strongly with each other (Fig. 4f).
IgM and IgG1 contribute most to SARS-CoV-2 neutralization
Given our observation that Ab isotype levels and neutralization titers varied tremendously among convalescent COVID-19 individuals (Figs. 2 and 5), we investigated the relative contribution of each Ab isotype to the neutralizing activities. Regression analyses were performed on 27 COVID-19-convalescent samples (TF#11 was excluded due to its outlier neutralization titer). As expected, relatively high r2 values (0.32 – 0.62) and significant p values were observed with total Ig, IgM and IgG1; in each case, r2 values were higher for spike than for RBD (Fig. 6a). For other isotypes, significant p values were sporadically achieved, but r2 values were weak (Fig. 6a,b).

Table showing sex (purple, F: female, M: male), relative levels of spike-specific (green) and RBD-specific (blue) Ig isotypes (+: bottom quartile, ++: second quartile, +++: third quartile, ++++: top quartile, -: non-responder) and reciprocal IC50 and IC90 neutralization titers against WT pseudovirus (orange) and D614G pseudovirus (red) of 29 plasma samples from COVID-19-convalescent individuals (nd: not done).

Simple linear regression of reciprocal IC90 neutralization titers of 27 COVID-19-convalescent individuals versus (a) spike-specific or (b) RBD-specific total Ig, IgM, IgG1 and IgA1 Ab levels. The black dash line shows 95% confidence interval. The dotted vertical red line represents the cut-off (mean of 12 pre-pandemic samples + 3 SD) for each isotype from Fig. 1. (b) Simple linear regression of reciprocal IC90 neutralization titers of 27 COVID-19-convalescent individuals versus spike-specific or RBD-specific total IgG2-4 and IgA2 Ab levels.
Neutralizing activities are mediated by IgM, IgG, and IgA fractions
To assess directly the capacity of different isotypes to mediate neutralization, we evaluated the neutralization activities of IgM, IgG, and IgA fractions purified from the plasma of five COVID-19-convalescent individuals (RP#1-5). The enrichment of IgM, IgG1, and IgA1 Abs reactive with spike and RBD was validated using the isotyping method used above (Supplementary Fig. 7 and not shown). These IgM, IgG, and IgA fractions were then evaluated for neutralizing activity along with the original plasma (Fig. 7). The RP#1-5 plasma neutralizing reciprocal IC50 titers ranged from 35 to 690 (Fig. 7a,b). Purified IgM and IgG fractions from RP#1-5 all mediated neutralization reaching more than 50%. Unexpectedly, plasma IgA fractions also displayed neutralizing activity, although not to the same potency as IgM and IgG (Fig 7c,d). In contrast, IgM, IgG, and IgA fractions from the negative control (RN#1) showed no neutralization (Fig. 7c,d).

(a) Neutralization of COV2pp by five COVID-19-infected individual plasma samples (RP#1-5) compared to a COVID-19-negative sample (RN#1). Plasma samples were tested at 4-fold dilutions from 1:10 to 1:40,960 or 1:20 to 1:81,920. Data are shown as the mean percentage of neutralization. The dotted horizontal lines highlight 50% neutralization. (b) Reciprocal IC50 and IC90 neutralization titers of RP#1-5 plasma samples (c) Neutralization of COV2pp by purified IgM, IgG, and IgA fractions from five COVID-19-infected individuals (RP#1-5) compared to a control Ig fraction. The fractions were tested at 4-fold dilutions from 500 to 0.02 µg/mL. Data are shown as the mean percentage of neutralization. The dotted horizontal lines highlight 50% neutralization. (d) IC50 of purified IgM, IgG, and IgA fractions from RP#1-5. The statistical significance was determined by a two-tailed Mann-Whitney test (*: p < 0.05, **: p < 0.01).
Discussion
Our study demonstrated that IgG1, IgA1 and IgM Abs against spike and RBD were highly prevalent in the plasma samples of convalescent COVID-19 patients approximately one to two months after infection. The presence of these isotypes was detected within 7-8 days after the onset of symptoms. Importantly, all three isotypes show the capacity to mediate virus neutralization. While regression analyses indicate the strongest contribution of IgM and IgG1 Abs to neutralizing activity, direct testing of purified isotype fractions showed that IgA also contributed to neutralizing activity, indicating the protective potential of all three major Ig isotypes. These data carry important implications for the use of convalescent plasma and hyperimmunoglobulin as COVID-19 therapeutic modalities, suggesting that the selection be based on the measurement of all of these Ig isotypes.
While all COVID-19 convalescent individuals exhibited plasma/serum neutralization activities reaching 50% neutralization, and 26 of 28 specimens attained 90% neutralization, neutralization levels were highly variable with reciprocal IC50 and IC90 titers ranging over three orders of magnitude. The titers were comparable against the initial Wuhan strain and the currently prevalent D614G strain. Similarly, the levels of spike- and RBD-binding total Ig and Ig isotypes varied greatly.
A trend toward higher levels of total Ig and each Ig isotype was seen in female compared to male subjects, as reported in another study56. Moreover, except for TF#11 (a male elite neutralizer), the median neutralizing reciprocal IC90 titer was higher in females than males, although the difference did not reach significance (data not shown). Sex differences in Ab induction have been observed following vaccination against influenza in humans and mice and were shown to result from the impact of sex steroids57,58. Whether and to what extent this contributes to sex differences seen in clinical outcomes of COVID-1959 remains to be investigated. Other studies have shown that the Ab levels were associated with multiple factors, including time from disease onset60 and disease severity30. However, other than sex, clinical data are not available for the subjects studied here, limiting our analysis only to neutralization and Ig isotypes.
One remarkable finding from our study is that although neutralization titers correlated with binding levels of IgM and IgG1 and not with those of IgA1 or IgA2, purified IgA fractions from convalescent COVID-19 patients exhibited significant neutralizing activities. The importance of this finding is underscored by the data showing that IgA1 was the prominent isotype in some plasma samples such as TF#7 and TF#24 and that IgA1 could be detected early, within a week after symptom onset. Data from other studies also supported IgA’s significance in that purified IgA fractions exhibited more or as potent neutralizing activities compared to purified IgG and that RBD-binding IgA correlated as strongly as the IgG equivalent with micro-neutralization titers61,62. The presence of IgA was also detected in the saliva and bronchoalveolar lavage samples from COVID-19 patients62,63. Nonetheless, Wang et al. reported that plasma IgA monomers were less potent than the plasma IgG and secretory IgA counterparts64. In our study, neutralization activities detected in the IgA fractions were mediated mainly by IgA1, the predominant IgA isotype in the plasma, and the IC50 potency of the IgA fraction was ∼4-fold lower than those of IgM and IgG1 fractions. This difference cannot be explained entirely by lower amounts of spike-specific IgA1 in the tested fractions, as estimations using spike-specific monoclonal antibodies of the respective isotypes yielded similar concentrations in IgA1 and IgM fractions (median of 2 and 2.5 µg/mL respectively). Fine epitope specificity and affinity may differ for IgA, IgM, and IgG to impact neutralization potency, but are yet to be evaluated.
In addition to neutralization, non-neutralizing Ab activities have been implicated in protection from virus infection through potent Fc-mediated functions such as antibody-dependent cellular cytotoxicity (ADCC), antibody-dependent cellular phagocytosis (ADCP), and complement-mediated lysis; this is reported for HIV, influenza, Marburg, and Ebola viruses44,65–68. The Fc activities were not evaluated in our study, and their contribution to protection against infection and disease progression in humans is yet unclear15,69–71. Interestingly, a recent study demonstrated enrichment of spike-specific IgM and IgA1 Abs and spike-specific phagocytic and antibody-dependent complement deposition (ADCD) activity in plasma of individuals who recovered from a SARS-CoV-2 infection, while nucleocapsid-specific IgM and IgA2 responses and nucleocapsid-specific ADCD activity were features enriched in deceased patients72. DNA vaccines expressing full-length and truncated spike proteins could curtail SARS-CoV-2 infection in the respiratory tract by varying degrees in rhesus macaques. Virus reduction correlated with levels of neutralization and also Fc-mediated effector functions such as ADCD15. Interestingly, these DNA vaccines elicited spike- and RBD-specific IgG1, IgG2, IgG3, IgA, and IgM Abs, and similar to our findings, neutralization correlated most strongly with IgM. Adenovirus serotype 26 vaccine vectors encoding seven different SARS-CoV-2 spike variants showed varying protection levels, and virus reduction correlated best with neutralizing Ab titers together with IgM binding levels, FcγRII-binding, and ADCD responses73. Defining the full potential of Abs against SARS-CoV-2 that includes neutralizing, non-neutralizing and enhancing activities are vital for determining the optimal convalescent Ab treatment against COVID-19 and assessing the potential efficacy of COVID-19 vaccine candidates.
When we examined plasma specimens collected within 7-8 days after COVID-19 symptom onset, we detected IgG and IgA against spike and RBD, as well as IgM. This is consistent with published reports showing that 100% of COVID-19-infected individuals developed IgG within 19 days after symptom onset and that seroconversion for IgG and IgM occurred simultaneously or sequentially30. IgA was also found early after infection (4-6 days after symptom onset) and increased over time in several other studies28,35,63,74. These studies suggest that measuring total Ig, rather than IgG, would provide a better outcome for early disease diagnosis. Indeed, we found no correlation between the levels of different isotypes examined in our study (data not shown). This lack of correlation may result from their asynchronous, sequential induction (IgM first, then IgG, and finally IgA). Still, IgA’s presence early during acute infection also suggests the potential contribution of natural IgA, which, similar to natural IgM, arises spontaneously from innate B1 cells to provide the initial humoral responses before the induction, maturation, and class-switching of adaptive classical B cells75,76.
In summary, this study demonstrates that spike- and RBD-specific IgM, IgG1, and IgA1 Abs were present in serum and plasma of all or almost all analyzed COVID-19 convalescent subjects and were detected at very early stages of infection. The plasma of convalescent individuals also displayed neutralization activities mediated by IgM, IgG, and IgA1, although neutralization titers correlated more strongly with IgM and IgG levels. The contribution of IgM, IgG, and IgA Abs to the neutralizing activities against SARS-CoV-2 demonstrates their importance in the efficacy of convalescent plasma used for COVID-19 treatment.
Methods
Recombinant proteins
The recombinant spike and RBD proteins were produced as previously described77 in Expi293F cells (ThermoFisher) by transfections of purified DNA using an ExpiFectamine Transfection Kit (ThermoFisher). The soluble version of the spike protein included the protein ectodomain (amino acids 1-1213), a C-terminal thrombin cleavage site, a T4 foldon trimerization domain, and a hexahistidine tag. The protein sequence was also modified to remove the polybasic cleavage site (RRAR to A) and two stabilizing mutations (K986P and V987P, wild type numbering). The RBD (amino acids 319-541) included the signal peptide (amino acids 1–14) and a hexahistidine tag. Supernatants from transfected cells were harvested three days after the transfection by centrifugation of the culture at 4000 g for 20 minutes. The supernatant was then incubated with 6 mL Ni-NTA agarose (Qiagen) for one to two hours at room temperature. Next, gravity-flow columns were used to collect the Ni-NTA agarose, and the protein was eluted. Each protein was concentrated in Amicon centrifugal units (EMD Millipore) and resuspended in phosphate buffered saline (PBS).
Human samples
Twenty-five citrated plasma of convalescent COVID-19 individuals destined for transfusion to SARS-CoV-2-infected individuals (TF#1-25, collected between March 26th and April 7th 2020) and ten specimens derived from the blood bank (N#4-13), representing contemporary COVID-19-negative blood bank donors, were obtained from the Division of Transfusion Medicine of the Department of Pathology, Molecular and Cell-Based Medicine (Mount Sinai Hospital System, IRB #20-03574). Four additional de-identified serum specimens from individuals with COVID-19 (P#5-8) were provided by the Clinical Pathology Division of the Department of Pathology, Molecular and Cell-Based Medicine at the Icahn School of Medicine at Mount Sinai. Serum and plasma samples were also obtained from study participants enrolled in IRB-approved protocols at the Icahn School of Medicine at Mount Sinai (Icahn School of Medicine at Mount Sinai IRB #16-00772, #16-00791, #17-01243) and the James J. Peter VA Medical Center (IRB #BAN-1604). Samples from these protocols included sera from seven participants with documented SARS-CoV-2 infection (P#1 d8, d11, and d15 after symptom onset, P#2 d7 and d10 after symptom onset, and RP#1-5 after convalescence), and sera from twelve healthy donors (N#1-3, N#14-22) collected prior to the spread of SARS-CoV-2 in the USA. All study participants provided written consent at enrollment and agreed to sample banking and future research use of their banked biospecimens. All samples were heat-inactivated and/or treated with 0.05% Triton X-100 prior to use.
Ig fractionation
IgA was first isolated from plasma by mixing 1:2 diluted plasma with peptide M agarose beads (600 µL/28 mL plasma, InvivoGen #GEL-PDM) for 1.5 hours at room temperature. Beads were then collected on a column and washed with PBS until protein reading (280 nm) by Nanodrop reached background. IgA was eluted from beads with a pH 2.8 elution buffer (Thermo Scientific #21004) and neutralized with pH 9 Tris buffer. The pass-through plasma sample was collected for IgG enrichment using protein G agarose beads (InvivoGen #GEL-AGG) using the same protocol as above and subsequently for IgM isolation using a HiTrap IgM column (G.E. Healthcare #17-5110-01) according to the manufacturer’s instruction. An additional purification step was performed using Protein A Plus mini-spin columns to separate IgG from IgM. Protein concentrations were determined with Nanodrop prior to use in Luminex and neutralization experiments.
Multiplex bead Ab binding assay
The SARS-CoV-2 antigens used in this assay consisted of a soluble recombinant trimerized form of the spike protein and a recombinant RBD protein78. Antigens were coupled to beads as previously described, with minor changes53. Each antigen was covalently coupled individually to a uniquely labeled fluorochrome carboxylated xMAP bead set at 2.0 μg protein/million beads using a two-step carbodiimide reaction with the xMAP Ab Coupling (AbC) Kit following to the manufacturers’ instructions (Luminex, Austin, TX). The coupled beads were pelleted, resuspended at 5×106 beads/mL in storage buffer (PBS, 0.1% bovine serum albumin (BSA), 0.02% Tween-20, and 0.05% sodium azide, pH 7.4), and stored at −80°C. Three to five million beads per batch were prepared in a 1.5 mL conical tube.
Before each experiment, the beads needed for a single run (2,500 beads/well x number of wells) were pelleted and resuspended in assay buffer (PBS, 0.1% B.S.A., 0.02% Tween-20) to deliver 2,500 beads in a volume of 50 μL/well in a 96-well plate. Sera/plasma samples were serially titrated (1:50 to 1:6400 final dilution) or diluted in assay buffer to 1:100 (for a final dilution of 1:200). The samples were then added as 50 μL/well to the wells containing the beads and incubated at room temperature for 1 hour at 600 rpm. After two washes in assay buffer, 100 μL/well of biotinylated antibodies specific for total Ig, IgG1, IgG2, IgG3, IgG4, IgA1, IgA2, or IgM was added and incubated for 30 minutes at room temperature on a plate shaker; these antibodies were rabbit biotinylated-anti-human total Ig (Abcam, catalog #ab97158) at 2 μg/mL, mouse biotinylated-anti-human IgG1 Fc (Invitrogen #MH1515) at 4 μg/mL, mouse biotinylated-anti-human IgG2 Fc (Southern Biotech #9060-08) at 1 μg/mL, mouse biotinylated-anti-human IgG3 Hinge (Southern Biotech #9210-08) at 3 μg/mL, mouse biotinylated-anti-human IgG4 Fc (Southern Biotech #9200-08) at 4 μg/mL, mouse biotinylated-anti-human IgA1 Fc (Southern Biotech #9130-08) at 4 μg/mL, mouse biotinylated-anti-human IgA2 Fc (Southern Biotech #9140-08) at 4 μg/mL or goat biotinylated-anti-human IgM (Southern Biotech #2020-08) at 3 μg/mL. After two washes, 100 μL/well of Streptavidin-Phycoerythrin (P.E.) at 1 μg/mL was added (BioLegend #405204) followed by a 30 minutes incubation at room temperature on a plate shaker. After two additional washes, 100 μL of assay buffer/well was added and put on a shaker to resuspend the beads. Each plate was read with a Luminex Flexmap 3D instrument. Specimens were tested in duplicate, and the results were recorded as mean fluorescent intensity (MFI).
COV2pp production and titration
The SARS-CoV-2 pseudoviruses (COV2pp) with wild-type (WT) or D614G mutated spike proteins were produced as previously described54. Briefly, 293T cells were transfected to overexpress SARS-CoV-2 glycoproteins. For background entry with particles lacking a viral surface glycoprotein, pCAGG empty vector was transfected into 293T cells. Around 8 hours post-transfection, cells were infected with the VSVΔG-rLuc reporter virus for 2 hours and then washed with PBS. Two days post-infection, supernatants were collected and clarified by centrifugation at 1250 rpm for 5 minutes. At the time of collection, a small batch of VSVΔG-rLuc particles bearing the CoV2pp was treated with TPCK-treated trypsin (Sigma-Aldrich #T1426-1G) at room temperature for 15 minutes prior to inhibition with soybean trypsin inhibitor (Fisher Scientific #1707502). Finally, particles were aliquoted prior to storage in −80°C.
The pseudoviruses were titrated on 20,000 Vero-CCL81 cells seeded in a 96-well black plate with clear bottom 24 hours before infection. At 18 to 22 hours post-infection, the infected cells were washed with PBS and processed for detection of Renilla luciferase activity with Renilla-Glo™ Luciferase Assay System (Promega #E2720). A Cytation3 (BioTek) instrument was used to read luminescence.
COV2pp neutralization
The day before infection, 20,000 Vero-CCL81 cells per well were seeded in a 96-well black plate with clear bottom. On the assay day, the COV2pp WT or D614G virus was diluted based on the titration results, and 82.5 µL added to all wells, except six for cell control, of a 96-well V-bottom plate. The seven serial dilutions of the samples were then prepared in another 96-well V-bottom plate. Samples of the COVID-19-infected individuals and COVID-19-negative individuals were tested at 4-fold dilutions from 1:10 to 1:40,960. Purified IgM, IgG, and IgA fractions and negative control fractions were tested at 4-fold dilutions from 500 to 0.02 µg/mL. 27.5 µL per well of the diluted sample was then added to the plate with the pseudovirus. For each plate, six wells were kept with the virus only, as virus control, and six wells with media only, as cell control. The plates were then incubated for 30 minutes at room temperature. 100 µL of the virus/sample mix or virus or medium alone was then added to the cells and spinoculated by centrifugation at 1250 rpm for 1 hour at room temperature. After incubation for 18 to 22 hours at 37°C, the measurement of infection/neutralization was performed as described for the COV2pp titration.
The percentage of neutralization was calculated with the formula: 100-((sample’s R.L.U. – cell control R.L.U.) / virus control R.L.U.) *100). The inhibitory concentration 50% (IC50) and 90% (IC90) were respectively defined as the reciprocal sample dilution or purified Ig fraction concentration achieving 50% and 90% neutralization.
Statistical analysis
Statistical tests that included two-tailed Mann-Whitney test, Spearman rank-order correlation test, and simple linear regressions were performed as designated in the figure legends. All statistical tests were performed using GraphPad Prism 8.
Data availability
The raw data that support the findings of this study are available from the corresponding author upon request.
Funding
This study was supported by the Microbiology Laboratory Clinical Services at the Mount Sinai Health System and the Mount Sinai Health System Translational Science Hub, National Institutes of Health [grant U54TR001433]; the Personalized Virology Initiative supported by institutional funds and philanthropic donations (to V.S.); the Department of Medicine of the Icahn School of Medicine at Mount Sinai Department of Medicine (to S.Z-P., C.E.H.); the Department of Microbiology and the Ward-Coleman estate for endowing the Ward-Coleman Chairs at the Icahn School of Medicine at Mount Sinai (to B.L.), the National Institute of Allergy and Infectious Diseases Centers of Excellence for Influenza Research and Surveillance [contract HHSN272201400008C] (to F.K., V.S.); the Department of Veterans Affairs [Merit Review Grant I01BX003860] (to C.E.H.) and [Research Career Scientist Award 1IK6BX004607] (to C.E.H.); the National Institute of Allergy and Infectious Diseases [grant AI139290] to C.E.H., [grant AI136916] (to V.S.), [grants R01 AI123449, R21 AI1498033] to B.L.
K.Y.O. and C.S. were supported by Viral-Host Pathogenesis Training Grant T32 AI07647; K.Y.O. was additionally supported by F31 AI154739. S.I. and C-T. H. were supported by postdoctoral fellowships from CHOT-SG (Fukuoka University, Japan) and the Ministry of Science and Technology (MOST, Taiwan), respectively.
Acknowledgments
We thank all the donors for their contribution to research.
Competing interests
The authors declare no competing interests.
Author contributions
J.K., S.W., G.E-A., S.Z-P., and C.E.H. wrote and edited the manuscript. S.W., J.K., C.E.H., and S.Z-P. designed the experiments. J.K., S.W., V.I., X.L. performed the experiments and collected the data. J.K., A.N., S.Z-P. and C.E.H. analyzed the data. K.Y.O., C.S., S.I., C-T.H., F.A., B.L., and F.K. provided protocols, antigens, cells and pseudovirus stocks. G.E-A., I.B., S.A., J.C.B., E.M.K., J.S., S.L., D.J., M.B-G., and V.S. provided samples. All authors read and approved the final manuscript.
Figure legends
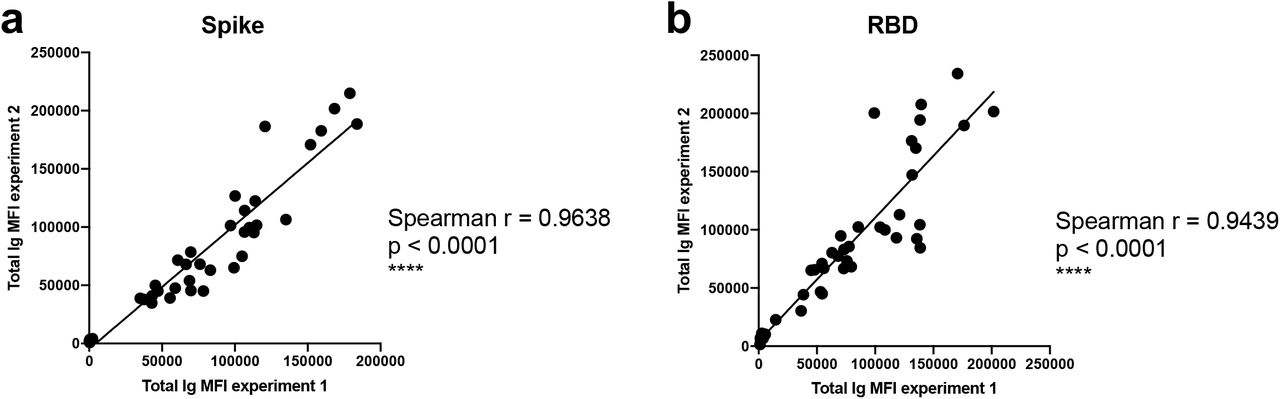
Spearman correlations of (a) spike-specific or (b) RBD-specific total Ig MFI values from two independent experiments to show the degree of assay reproducibility.

Spearman correlations of the area under the curves (AUCs) of (a) spike- or (b) RBD-specific total Ig versus total Ig MFI values at a 1:200 dilution.
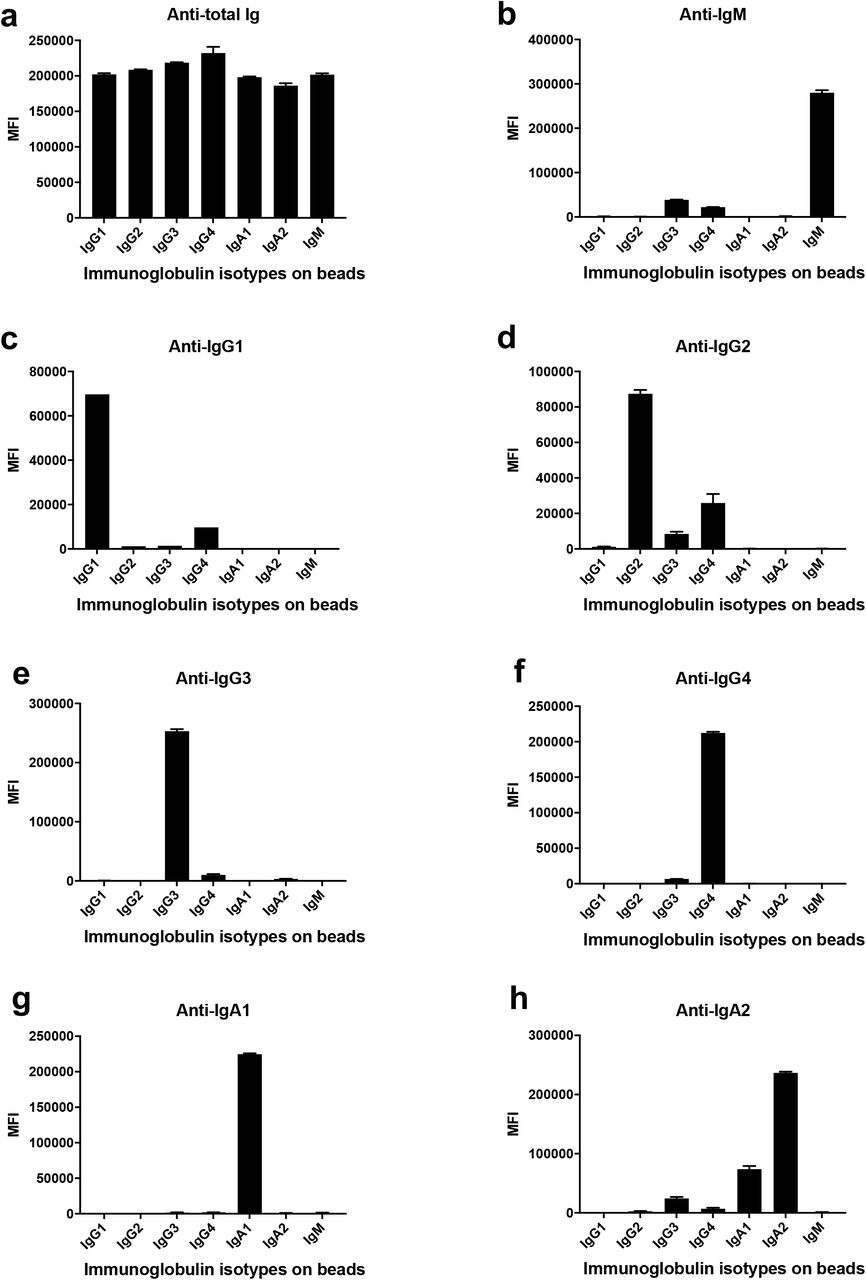
Isotyping validation was performed by coating Luminex beads with IgG1, IgG2, IgG3, IgG4, IgA1, IgA2, and IgM myeloma proteins and detecting each specifically with eight different secondary Abs against (a) total Ig, (b) IgM, (c) IgG1, (d) IgG2, (e) IgG3, (f) IgG4, (g) IgA1 and (h) IgA2. The data are shown as mean MFI + SD of duplicate.

Spearman correlations between spike-specific versus RBD-specific total Ig, IgM, IgG1, IgG2, IgG3, IgG4, IgA1, or IgA2 MFI values.

Violin plots of (a) spike-specific or (b) RBD-specific total Ig, IgM, IgG1, and IgA1 levels from nine COVID-19 convalescent female (F) and 15 male (M) subjects. The statistical significance was determined by a two-tailed Mann-Whitney test (ns: non-significant: p > 0.05).
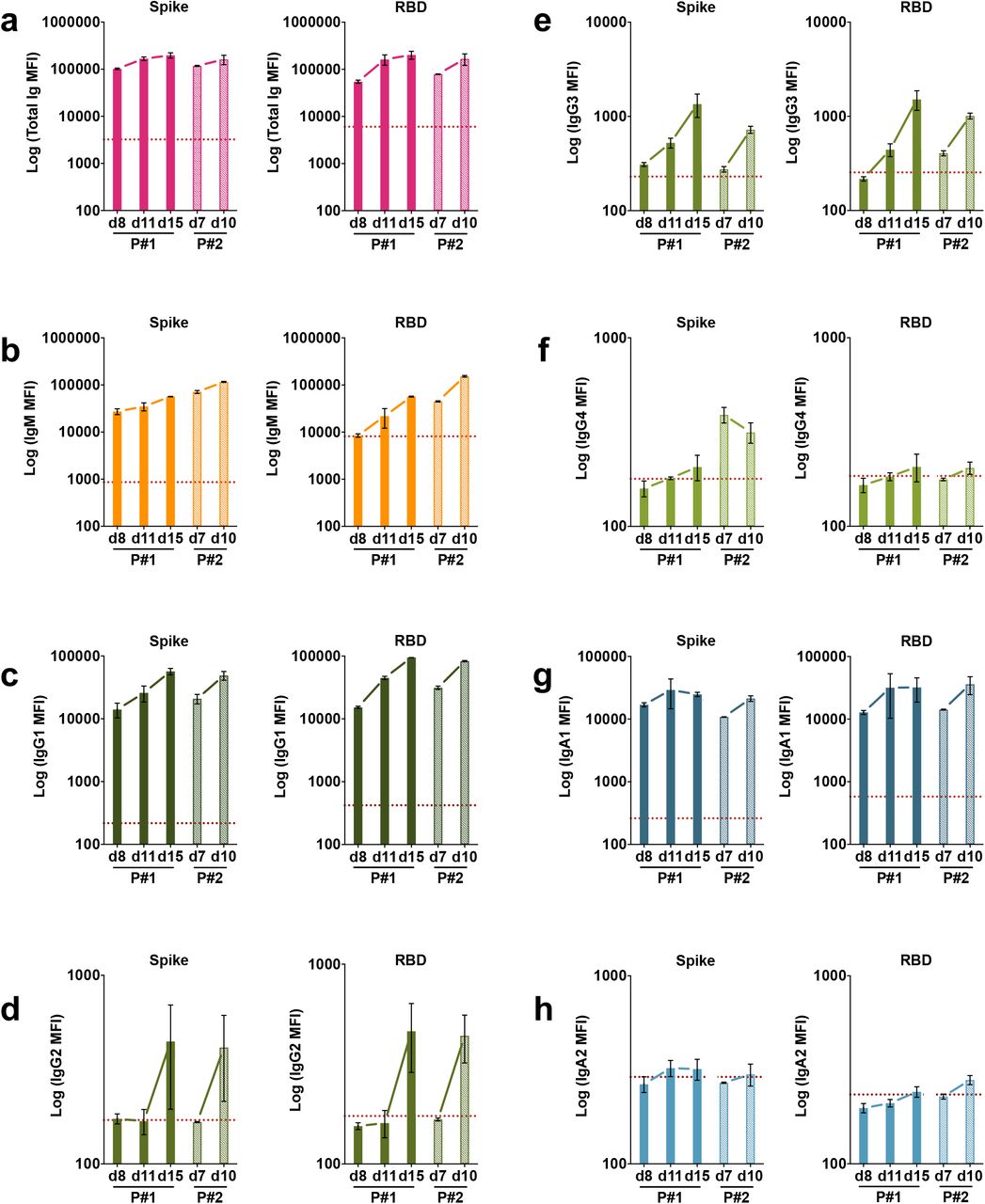
Kinetics of induction of spike-specific (left panel) or RBD-specific (right panel) (a) total Ig, (b) IgM, (c) IgG1, (d) IgG2, (e) IgG3, (f) IgG4, (g) IgA1, and (h) IgA2 from two COVID-19 patients. Longitudinal samples from each patient were tested at a dilution of 1:200 in parallel with all negative samples and data are shown as mean MFI + SD of duplicate measurements from at least two experiments. The dotted red line represents the cut-off value calculated as the mean of 12 pre-pandemic samples + 3 SD from Fig. 1.
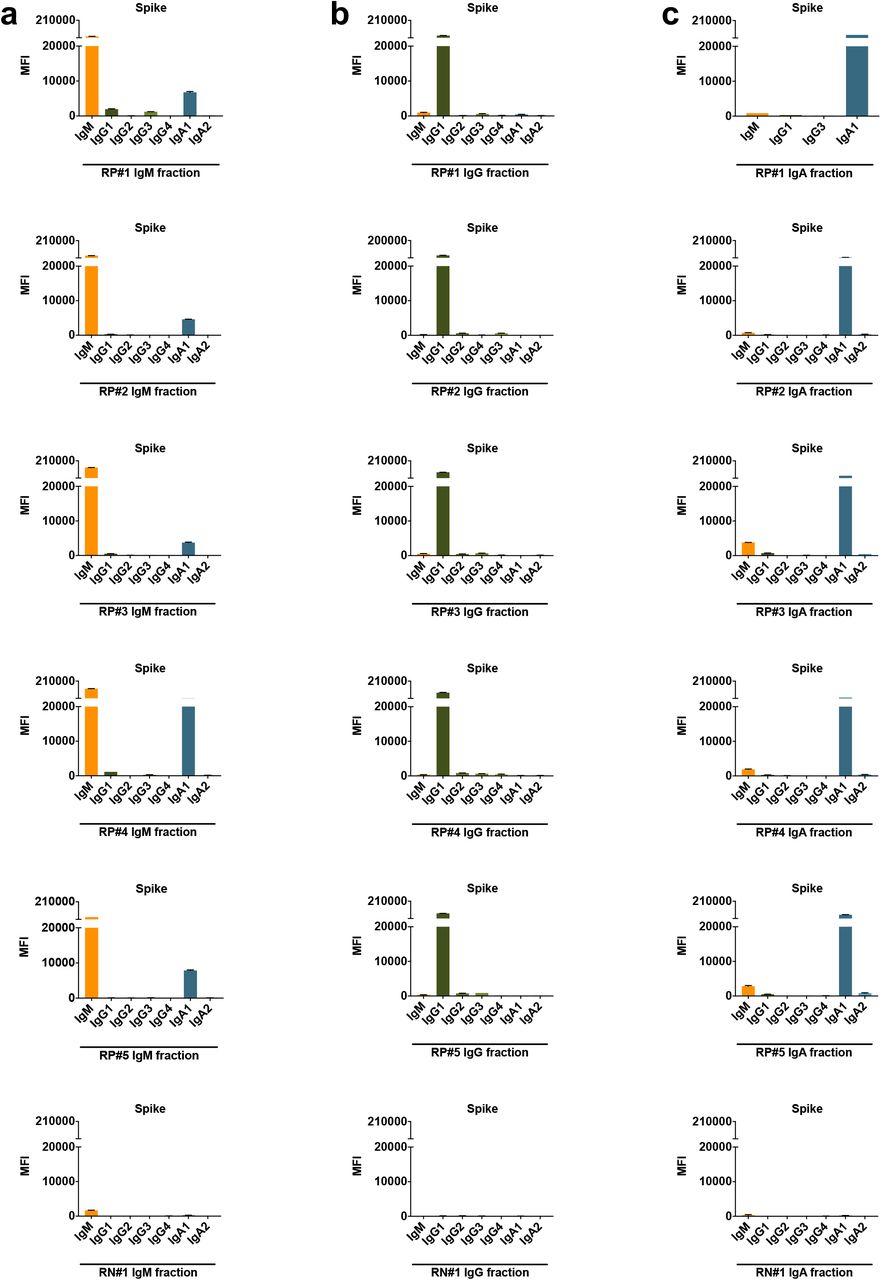
Each fraction was measured for the presence of IgM, IgG1, IgG2, IgG3, IgG4, IgA1, and IgA2 Abs using the isotyping method validated in Supplementary Fig. 3.
Footnotes
Pre-pandemic controls included, neutralization of the D614 mutant added, statistical analyses updated, a co-author added.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}