
Abstract
Aldehydes are toxic chemicals rapidly metabolized by the enzyme aldehyde dehydrogenase 2 (ALDH2). Nearly 8% of the world population carry an ALDH2 genetic variant rs671 (E504K) causing inefficient aldehyde metabolism recognized by facial flushing after alcohol consumption. Importantly, rs671 carriers have a several-fold increased risk of cancer, neurodegenerative, and heart disease attributed to inefficient aldehyde metabolism. Although rs671 is common, it is unlikely rs671 is the only inactivating ALDH2 mutation in humans.
By recruiting human volunteers reporting facial flushing after alcohol consumption, we identified previously uncharacterized ALDH2 point mutations including rs747096195 (R101G), rs190764869 (R114W), and rs4646777 (G>A). Using ALDH2 recombinant enzyme, we determined R101G and R114W have a reduced efficiency in acetaldehyde metabolism unique to E504K (Vmax/Km, 1.95±0.01+ and 2.77±0.02+ relative to 0.93±0.03, respectively n=3/group, +P<0.001 relative to 7.97±0.51 for wild type ALDH2).
When subjecting human volunteers to an alcohol challenge (0.25g/kg), acetaldehyde levels increased 9-fold for rs671 (E504K), 2-fold for rs747096195 (R101G) and rs190764869 (R114W) and was no different for rs4646777 (G>A) relative to volunteers carrying wild type ALDH2. Further, for volunteers tested, heart rate changes after alcohol strongly correlated with decreased acetaldehyde metabolism (r=0.90, n=26) as opposed to facial flushing. Taken together, we identified novel inactivating ALDH2 point mutations and developed a non-invasive diagnostic test to phenotype aldehyde metabolism in humans. Identifying ALDH2 genetic differences and their phenotype in response to alcohol can provide more precise individual recommendations regarding the health risks of aldehyde exposure.
Introduction
Aldehydes are toxic compounds that can be inhaled, consumed, or endogenously produced1-4. Exogenous exposures of aldehydes, such as acetaldehyde and formaldehyde, can occur from sources such as air pollution, cigarette smoke, and alcohol2. Endogenous sources of aldehydes, such as 4-hydroxynonenal and malondialdehyde, are produced from lipid peroxidation such as during surgery or organ injury3, 4. As aldehydes are highly diffusible and penetrate membranes, aldehydes intracellularly can cause aldehyde-induced DNA and protein adducts that are mutagenic, carcinogenic, and cytotoxic which in turn drive the pathogenesis for cancer, cardiovascular, and neurodegenerative diseases5, 6.
A major protective mechanism the cell has against aldehyde-induced toxicity is the enzyme aldehyde dehydrogenase 2 (ALDH2) which acts to neutralize and limit aldehyde-induced damage3. However, nearly 540 million people world-wide, mainly of East Asian descent, carry an ALDH2 genetic variant rs671 (E504K) decreasing ALDH2 enzymatic activity by at least 50% relative to the wild type ALDH2 enzyme7. With aldehyde exposures, rs671 is less efficient in metabolizing aldehydes. Subsequently, aldehyde accumulation and intracellular aldehyde-induced adduct formation occurs in greater frequency relative to those carrying the wild type ALDH2 enzyme; leading to higher risks of cancer, cardiovascular, and neurodegenerative diseases for people carrying rs6714, 8-10.
Although rs671 is common, it is unlikely the only inactivating ALDH2 mutation. In humans, several additional ALDH2 missense mutations, including a common intron mutation (rs4646777, G>A, 0.18 frequency), are identified by whole genome sequencing11. However, whether these additional ALDH2 mutations are inactivating mutations are unknown. As such, screening ALDH2 mutations for those causing inefficient aldehyde metabolism is important considering the health risks associated with the rs671 variant and aldehyde exposure12.
Facial flushing after alcohol can be an indicator of aldehyde accumulation. For those carrying rs671, the acetaldehyde accumulation after alcohol consumption produces facial flushing and tachycardia8; a condition called Alcohol Flush Syndrome13. However, it is also unlikely that all people who have facial flushing after alcohol accumulate acetaldehyde. Thus, a means to identify people who accumulate aldehydes due to carrying an ALDH2 point mutation is important considering the health risks associated with rs671. Here we recruited human volunteers self-reporting facial flushing after alcohol consumption, characterized ALDH2 mutations identified using recombinant protein, and developed a non-invasive screening method to phenotype aldehyde metabolism in humans through using breath monitoring and physiological measurements.
Methods
Prior to recruitment and testing, IRB approval was obtained from Stanford University (IRB 46095). During IRB review, this study was identified as a basic experimental study involving humans with the manipulation or task used (consuming alcohol) expressly used for measurement and was not considered an intervention. Written informed consent was received from participants prior to inclusion in the study. The experiments conformed to the principles set out in the WMA Declaration of Helsinki and the Department of Health and Human Services Belmont Report.
Participant recruitment
Participants were recruited from December 2018 through March 2020 using flyers posted on the Stanford University campus in succession. The initial flyer recruited people who flush after they consume alcohol for an alcohol study. During the recruitment phase, public support for this project triggered the recruitment flyer at Stanford University to be posted on the Facebook group page Subtle Asian Traits that has ∼1.8 million members. Due to the public response caused by the flyer posted on a Facebook group page, the flyer was modified to recruit additional people only of non-Asian descent.
Participants were initially screened for exclusion using a questionnaire (see Supplemental Material which describes exclusion criteria). Participants were also instructed to complete a CAGE questionnaire to screen out participants with potential alcoholism. Those participants eligible then provided a saliva sample for DNA extraction and purification (Quick-DNA Miniprep Kit, Zymo Research). DNA was amplified using 12 primers designed to cover the ALDH2 exome for sequencing while in addition an ALDH2 common intron mutation (rs4646777) as described (Supplemental Table 1). After amplification, samples were sent for Sanger sequencing (Eton Bioscience).
In Silico ALDH2 Modeling
Protein data bank (PDB) entry 1O02 was imported into PyMOL (Schrödinger, Inc.) for molecular visualization of wild-type ALDH2 and to mutate R101 and R114 by using the protein mutagenesis function.
ALDH2 Site-Directed Mutant Construction, Protein Expression and Purification
Based upon genotyping results, site-directed mutagenesis of human ALDH2 was performed using a QuikChange site-directed mutagenesis kit (Agilent Technologies) using the kanamycin resistance template plasmid, pET-28a-c (+). Oligonucleotides used to generate ALDH2 mutants (R101G, R114W and E504K) were purchased (Integrated DNA Technologies). Co-expression of wild-type ALDH2 and its site directed mutants (R101G, R114W and E504K) was achieved using the pEDuet-1 vector plasmid (Millipore Sigma, Milwaukee, WI). One copy of wild-type ALDH2 was cloned into the His-tag cloning site of the pETDuet-1 vector plasmid by EcoRI and HindIII, whereas another copy was cloned into the S-tag using NdeI and XhoI. For co-expression of wild-type ALDH2 with the variants, one copy of wild type was cloned into the His-tag cloning site as described above, whereas the variants were cloned into the S-tag cloning site of pETDuet-1 plasmid using NdeI and XhoI. Co-expression of the recombinant proteins was carried out at 30 °C in the presence of an ALDH2 chaperone using E. coli BL21 (DE3) host cells. 0.5 M IPTG was used for induction at 30°C for 16 hours. Purification of the recombinant proteins was achieved using the His-trap nickel affinity column (GE Healthcare).
ALDH2 Enzymatic Activity Assay
NAD+ conversion to NADH in the presence of acetaldehyde was used to determine ALDH2 enzyme activity as previously described 14. A blank control was run without acetaldehyde. Results were collected in triplicates on two separate days.
Human Alcohol Challenge
Participants were selected for the alcohol challenge portion of the study based on genotyping results. At this stage, participants were also excluded if homozygous for rs671 (secondary to the expected severity of the alcohol response). Participants heterozygous for rs671 (E504K) were called back for participation in the alcohol challenge in addition to those without sequenced mutations were selected to closely match the age, sex and weight of the participants heterozygous for rs671 (E504K). Additional participants were selected for the alcohol challenge study based upon genotyping results identifying point mutations including rs747096195 (R101G), rs190764869 (R114W), and rs4646777 (G>A, intron 8).
Participants were instructed prior to the alcohol challenge to not eat or drink for 2 hours prior to testing. Testing was performed between 12pm and 4pm on a weekday. Upon arrival, patients were consented and weighed. To assess physiologic measurements, a five lead EKG was placed and EKG measurements were recorded (ADInstruments). Blood pressure and heart rate was measured using a standard blood pressure cuff and pulse oximeter (DynaMap). Skin temperature was measured under the left cheek bone using liquid crystal technology (Crystaline II Temperature Indicators, Sharn Anesthesia). Baseline hemodynamic data including blood pressure, heart rate, and skin temperature were collected. Prior to testing, a hand-held alcohol breath test meter confirmed no alcohol was consumed before testing (BACtrack Element Pro). Breath samples were collected by a 3L Tedlar bag (Zefon International) that were sealed after collection. Breath samples were analyzed within 4 hours of sample collection.
After establishing a baseline, participants were given a 415 mL (14oz) drink consisting of 0.25g/kg Ketel One vodka, water, and Minute Maid lemonade. For reference in the United States, a standard drink is 14 grams of alcohol which is equal to one 12 oz glass of 5% beer or one 5oz glass of 12% wine 15. As an example, a 70kg person in this study drank 17.5g of alcohol. Participants consumed the first 7oz of the 14oz drink during the first 5 minutes of the study and the second 7oz during the second 5 minutes. After baseline measurements taken 3 minutes before alcohol consumption, physiologic measurements of heart rate, blood pressure, skin temperature, and breath metabolites were taken at 5, 10, 15, 30, 60, 90, 120, and 150 minutes during and after alcohol consumption (Figure 2A). After completion of the study, a breath alcohol test and a standard field sobriety test was administered. The study lasted a total duration of 3 hours. When collecting the data, we considered the heart rate, blood pressure and facial skin temperature as the physiological measurements to analyze in relation to the acetaldehyde levels measured. Participants completing the alcohol challenge were given $125 compensation.

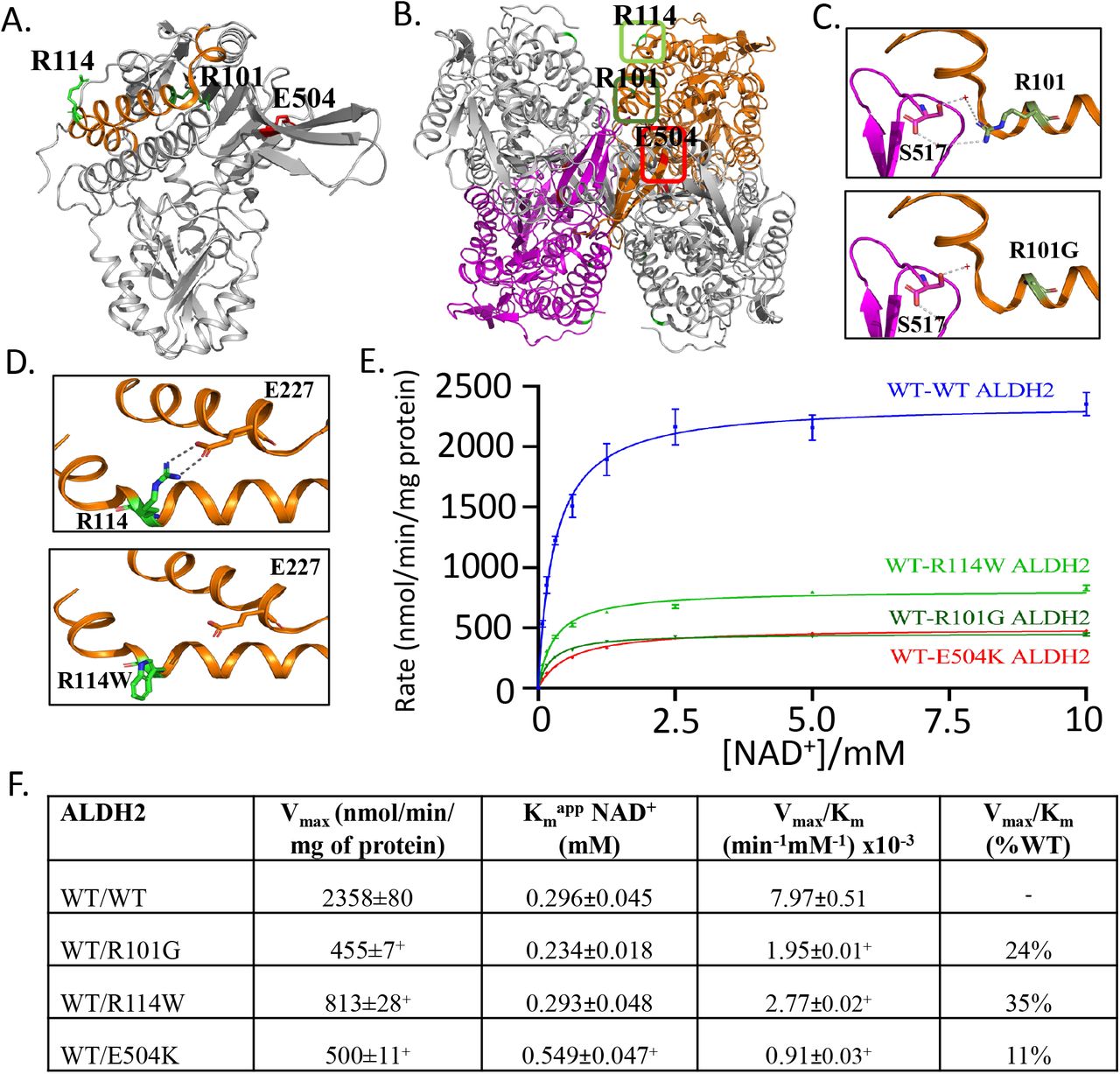
A. Crystal structure of ALDH2, with 3 amino acids within the structure highlighted, R101G (rs747096195, dark green), R114W (rs190764869, light green) and E504K (rs671, red). B. ALDH2 tetramer with location of R101 (dark green box) and R114 (light green box). C. R101G point mutation results in loss of a hydrogen bond involved in stabilizing ALDH2 dimerization. D. R114W point mutation results in loss of hydrogen bonds between the adjacent alpha helix within the ALDH2 monomer. E. In vitro results supporting inefficient acetaldehyde metabolism for R101G and R114W. F. Summary table of Vmax and Km for missense mutations R101G and R114W relative to wild type ALDH2 and E504K. n=3/group, +P<0.01 versus wild type ALDH2 enzyme
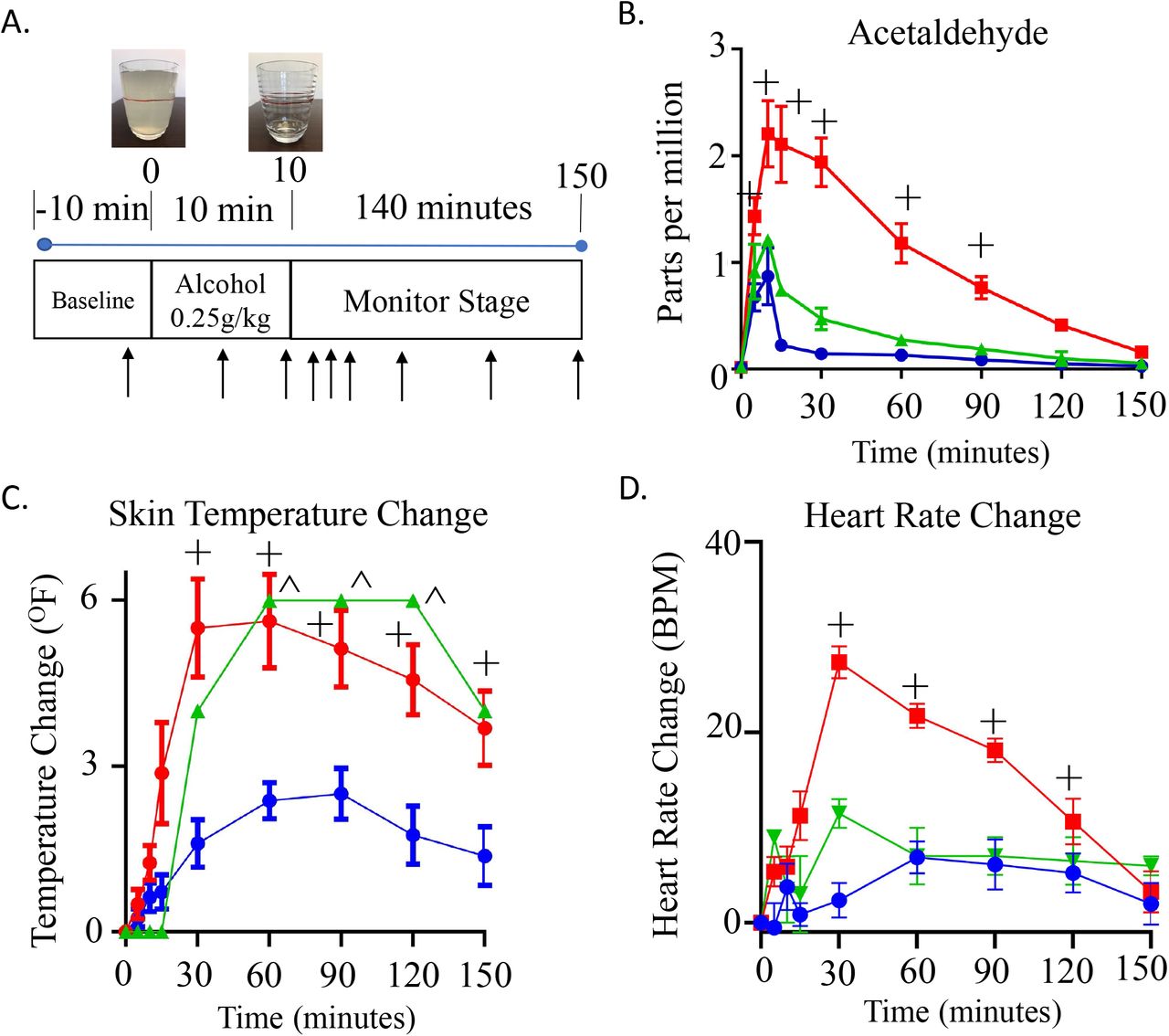
A. Alcohol challenge protocol. After 10 minutes baseline, subjects were given an alcohol challenge (0.25g/kg); consumed over 10 minutes. Subjects were monitored by measuring physiologic parameters. Breath measurements were measured at 9 time points (black arrow). After collecting a baseline measurement, the next 2 measurements were during alcohol consumption followed by monitoring for an additional 140 minutes after alcohol consumption. B. Breath concentration (in parts per million) of acetaldehyde. C. Change in skin temperature after alcohol consumption. D. Change in heart rate after alcohol consumption. For all panels, blue lines are wild type ALDH2 subjects (n=8), red lines are subjects that are heterozygotes for rs671 (E504K, n=8), green are heterozygotes for either rs190764869 (R101G) or rs747096195 (R114W). + or ^P<0.01 versus wild type ALDH2 subjects.
Breath metabolite analysis
The breath metabolite acetaldehyde was measured from a Tedlar bag using selective ion flow mass spectrometry within 4 hours of collection (Syft Voice Ultra, New Zealand). Soft ionization was carried out using H3O+ and NO+ with the mass and reaction ratios described for measuring acetaldehyde (Supplemental Table 2). Helium was used a carrier gas.
Statistical Analysis
All data was presented as mean ± SEM. The number needed to recruit for this study was based upon a prior study examining differences in acetaldehyde metabolism in the blood. Based upon the dose of alcohol (0.25g/kg) producing a 5-fold difference in blood acetaldehyde levels, we predicted that at a minimum a need to recruit 4 people per group for statistical differences between people that are wild type ALDH2 relative to people that are heterozygous for rs671 (E504K)16. For analysis of differences in metabolites or physiological data, a two-way ANOVA followed by Bonferroni correction for multiplicity was used in order to compare each group to the control group or to compare measurements performed within a group over time. When comparing heart rate changes with acetaldehyde levels, a Pearson correlation test with a 2-tailed p-value was used. Statistical analysis was performed using GraphPad Prism 6. + or ^=p < 0.01 was considered statistically significant between groups.
Results
Initially, 51 people participated in a phone screen, 29 people provided a saliva sample for genotyping, and 16 people were given an alcohol challenge where 8 were heterozygotes for rs671 and 8 were not (Supplemental Figure 1A). After the recruitment flyer was modified to recruit participants of non-Asian descent, 68 additional people underwent a phone screen, 37 people provided saliva samples, and based on the genotyping results, 10 people were given an alcohol challenge (Supplemental Figure 1B).
After genotyping, we identified people heterozygous for rs671 (E504K), rs747096195 (R101G), rs190764869 (R114W) and an intron rs4646777 (G>A). When examining the crystal structure, R101G and R114W are located in distinct regions of the α-helix that are separate from E504K (Figure 1A). A closer look at the ALDH2 tetramer suggests that R101 is important in the interaction between ALDH2 subunits while R114 is important in the interaction of alpha helices within the same subunit (Figure 1B). When R101 is mutated to a glycine, a hydrogen bond via a conserved water molecule is lost between R101 and S517 (Figure 1C). Upon R114W mutation, the two hydrogen bonds between E227, present within the same subunit on an adjoining alpha helix, are lost (Figure 1D). To mimic the heterozygous condition in humans, we co-expressed wild type and the ALDH2 point mutations into the pETDuet-1 vector. The co-expressed mutants R101G, R114W, and E504K had a significant decrease in Vmax compared to wild-type ALDH2 (Figure 1E and 1F, 455±7+, 813±28+, and 500±11+ versus 2358±80 nmol/min/mg protein for wild type ALDH2, respectively, n=3/group). Further, the Km for R101G and R114W co-expressed with wild-type ALDH2 were similar to wild type ALDH2 (Figure 1E and 1F, Km = 0.234±0.018 and 0.293±0.045 versus 0.296±0.045mM, respectively). However, E504K co-expressed with wild type ALDH2 had a ∼2-fold increase of Km compared to wild type, R101G and R114W ALDH2 (Figure 1F).
To explore the phenotype based on the ALDH2 genotyping from human subjects, we subjected participants who were heterozygous for rs747096195 (R101G), rs190764869 (R114W) and rs671 (E504K) to an alcohol challenge and compared the response to those participants carrying wild type ALDH2. We considered the rs747096195 and rs190764869 point mutations together as these point mutations are within the same alpha helix for ALDH2, each are in extremely low frequency in humans (0.000024 for rs747096195 and 0.0000121 for rs190764869)11, and have a similar Km based on in vitro data. For participants given an alcohol challenge, general demographics including age, weight, and sex were collected (Supplemental Table 3). When comparing heterozygotes for rs671 (E504K, ALDH2*1/*2) with respect to those with a wild type ALDH2 genotype, breath acetaldehyde levels peaked 9-fold 5 minutes after completing alcohol consumption (Figure 2B, 15-minute time point measurement, peak: 2.1±0.4+ versus 0.2±0.3 ppm, n=8/group). Further, rs747096195 (R101G) and rs190764869 (R114W) accumulated acetaldehyde and took longer to metabolize acetaldehyde relative to those subjects that were wild type ALDH2; supporting the recombinant enzyme studies. Additionally, skin temperature changes relative to baseline significantly increased for rs671 at 30 minutes followed by rs747096195 (R101G) and rs190764869 (R114W) at 60 minutes after the start of alcohol consumption (Figure 2C). Concomitantly, heart rate changes also markedly increased in rs671 (E504K) participants in addition to less substantial heart rate changes for those carrying rs747096195 (R101G) and rs190764869 (R114W) (Figure 2D).
Next, we tested additional participants that carried an intron mutation rs4646777 (Figure 3A, G>A, intron 8, 0.18 frequency) or mixed-race heterozygotes for rs671 (E504K). General demographics including age, weight, and sex are described (Supplemental Table 4). When subjected to an alcohol challenge, rs4646777 had minimal changes in acetaldehyde concentrations (Figure 3B), with metabolism nearly identical to those genotyped as wild type ALDH2 (Figure 2B). However, unlike those genotyped as wild type ALDH2, noticeable facial flushing with increases in facial skin temperature occurred after an alcohol challenge for participants either carrying rs4646777 or rs671 (Figure 3C). The timing of facial flushing was also different between rs4646777 or rs671 with rs4646777 having a more delayed facial flushing response that peaked 90 minutes after alcohol consumption. Heart rate changes after alcohol also occurred for those carrying rs671 as opposed to rs4646777 (Figure 3D).

{kind=link}
{kind=link}
{kind=link}
A. Intron location for rs4646777 B. Acetaldehyde levels after alcohol consumption for subjects heterozygotic for rs4646777 and subjects that were mixed-race for rs671. C. Skin temperature changes for rs4646777 and rs671. D. Heart rate changes for rs4646777 and rs671. E. Cumulative acetaldehyde levels after alcohol consumption for each group tested. Data is expressed as area under the curve (ppm-min). WT= wild type ALDH2. F. Heart rate change versus acetaldehyde levels at 30 minutes after starting alcohol consumption. rs671 (red and dark red), rs4646777 (grey), rs747096195 and rs190764869 (green), and wild type ALDH2 (blue). ˅P<0.01 relative to rs4646777, + or ^P<0.01 relative to wild type ALDH2 subjects.
Together, when comparing total acetaldehyde accumulation after an alcohol challenge for all subjects tested, acetaldehyde accumulation was 6-fold greater for heterozygotes with rs671 and more variable versus those without ALDH2 mutations (Figure 3E, 143±14+ versus 22±3 ppm-minutes). Further, total acetaldehyde was minimal for people with rs4646777 (20±2 ppm-minutes) as opposed to a two-fold accumulation for rs747096195 and rs190764869 (46±7 ppm-minutes) (Figure 3E). Heart rate changes 30 minutes after the start of alcohol consumption correlated with total acetaldehyde area under the curve (r=0.88, 95% confidence 0.74-0.94, R2=0.77, Supplemental Figure 3A) and acetaldehyde levels measured at 30 minutes after the start of alcohol consumption (r=0.90, 95% confidence 0.78-0.95, R2=0.80, Figure 3F). This was opposed to skin temperature changes that had less of a correlation with total acetaldehyde area under the curve (r=0.53, 95% confidence 0.19-0.76, R2=0.28, p<0.001, Supplemental Figure 3B) and acetaldehyde levels measured at 30 minutes after the start of alcohol consumption (r=0.55, 95% confidence 0.21-0.77, R2=0.30, Supplemental Figure 3C).
Discussion
Here we identified and characterized human ALDH2 point mutations using recombinant enzyme and developed a non-invasive method to screen for inactivating ALDH2 point mutations in humans. As we find here, additional inactivating ALDH2 mutations besides rs671 exist in humans; indicating greater than 8% of the world population are impacted by inefficient aldehyde metabolism. Importantly, people self-identifying as non-Asian also carry ALDH2 inactivating point mutations and ALDH2 inactivating point mutations are not exclusive to the Asian population. Therefore, screening for inactivating ALDH2 point mutations should be inclusive for all ethnic populations and with this approach can lead to developing precision medicine pathways that can provide more individualized recommendations regarding the health risks with aldehyde exposures. This is imperative since prior studies clearly support people carrying inactivating ALDH2 mutations, such as rs671, are at higher risk for cancer, cardiovascular, and neurological diseases with aldehyde exposure4.
Although facial flushing is commonly considered a sign for carrying an ALDH2 inactivating point mutation, our study does not support this association. Instead, monitoring heart rate changes after alcohol consumption can provide more detailed information regarding aldehyde metabolism beyond that of genetic information alone. Humans heterozygous for rs671 are non-homogenous in their response to aldehyde metabolism; stemming from ALDH2 enzyme activity being highly dependent on the ratio between E504K and E504 monomers forming the ALDH2 tetramer3. Therefore, assessing heart rate changes after alcohol, as we show here, can further categorize the severity of acetaldehyde accumulation from carrying an inactive ALDH2 mutation individually. Heart rate monitoring during alcohol consumption could also open up an opportunity for people to screen for inactive ALDH2 point mutations in their home by measuring heart rate before and after alcohol consumption by palpation or with a smartwatch.
This is also the first study to characterize and phenotype the novel ALDH2 point mutations rs747096195 (R101G) and rs190764869 (R114W). This inefficient aldehyde metabolism is different mechanistically relative to rs671 (E504K) as R101 and R114 are residues in a long α-helix that is in a distinctly different location to E50417. Recombinant enzyme activity of R101G and R114W co-expressed with wild-type ALDH2 demonstrated that the missense ALDH2 mutations have less activity (Vmax/Km) relative to wild-type ALDH2. In humans, acetaldehyde metabolism for rs747096195 (R101G) and rs190764869 (R114W) is reduced 2-fold rather than the 6-fold response for those carrying rs671 (E504K); likely since the Km for the ALDH2 enzyme is left unaffected by the R101G and R114W point mutations. Further, several other missense mutations exist within the exome for the ALDH2 enzyme12 and these ALDH2 mutations will have to be studied to understand the phenotype occurring in response to aldehyde exposure.
Additionally, rs4646777 causes facial flushing without acetaldehyde accumulation. The pathophysiological significance of rs4646777 is less known. One GWAS study involving a case-control study in a Norwegian population for lung cancer with 335 cases and 415 controls examining 105 SNPs identified a possible association of rs4646777 with lung cancer which later in this study was considered a false positive and could not be validated18. As GWAS studies have not identified rs4646777 as a potential cause of human pathophysiology, rs4646777 may cause facial flushing but potentially be benign. However, as rs4646777 has a high frequency (.18) relative to other ALDH2 mutations, it is important to further understand if there are health implications for carrying rs4646777 besides causing facial flushing after alcohol consumption.
In the past, rs671 carriers were considered protected from alcohol-related health problems by abstaining from alcohol use. However, even moderate alcohol consumption for rs671 carriers increases the odds ratio for developing aerodigestive track cancer (2.61, 1.19-5.75) and esophageal cancer (3.12, 1.95-5.01)19, 20. This is important to recognize as societal pressures occurring globally are steering people with ALDH2 inactivating mutations towards alcohol consumption. For example, although the prevalence of rs671 is 20% in the Republic of Korea, the work culture expects employees to attend dinners and drink alcohol. This culture likely fuels the Republic of Korea having the highest annual per capita alcohol consumption of 12.3L in the Asia Pacific region21. In East Asian countries, there is also a concerning trend that alcohol use disorder is steadily rising in heterozygotes who carry rs67122, 23. In the United States, social drinking and binge drinking at college is also concerning and may pressure those with ALDH2 inactivating mutations to drink alcohol24, 25. The COVID-19 pandemic has also seen a ∼27% increase in off-premise alcohol sales in the United States for April 2020 relative to the previous year26. Together, these social factors driving alcohol use need to be highlighted to raise public awareness for the importance of identifying inactivating ALDH2 point mutations that limit aldehyde metabolism when considering the carcinogenic effects of alcohol.
Our results need to be interpreted within the realm of potential limitations. As the recruitment flyers were designed to identify people that have facial flushing after they consume alcohol, it is untested whether aldehyde accumulation may occur without facial flushing. Furthermore, point mutations other than rs671 and rs4646777 are relatively rare in the general population, which prevented the collection of a large sample size for rs747096195 (R101G) and rs190764869 (R114W) that we identified. Regardless, the non-invasive assay we developed can provide individualized assessments of acetaldehyde metabolism; leading to more personalized medicine strategies by identifying those that have impaired acetaldehyde metabolism after alcohol consumption versus those that do not.
In summary, we identified novel point mutations in ALDH2 and developed a methodology to phenotype their response to alcohol. This can present an opportunity to develop precision medicine pathways and individualized recommendations for alcohol use to ultimately reduce alcohol-associated cancer risk.
Data Availability
All data is presented within the manuscript and is freely available.
Conflicts of Interest
Funding for this project was supported by Stanford ChEM-H (ERG and DMR), NIAAA AA011147 (DMR), and NIGMS GM119522 (ERG).
Eric Gross holds a patent related to the ALDH2 activator Alda-1 and research funding from the National Institutes of Health. Daria Mochly-Rosen and Che-Hong Chen hold patents related to Alda-1 and research funding from the National Institutes of Health. Dr. Angst has funding for a research study from Alkahest Inc. Additional authors have no disclosures.
Acknowledgements
The authors would like to thank Martina Steffen for her initial support as a clinical coordinator for this study and Kayla Anchondo-Johnson, RN for her assistance in collecting physiologic data.
References




